青藏高原木孜塔格冰川、玉珠峰冰川及扎当冰川可培养细菌的生理特征
【类型】期刊
【作者】邢婷婷,刘勇勤,王宁练,沈亮,顾政权,郭泌汐(中国科学院青藏高原研究所高寒生态学与生物多样性重点实验室;中国科学院青藏高原地球科学卓越中心;中国科学院青藏高原研究所青藏高原环境变化与地表过程重点实验室;中国科学院寒区旱区环境与工程研究所冰冻圈科学国家重点实验室)
【作者单位】中国科学院青藏高原研究所高寒生态学与生物多样性重点实验室;中国科学院青藏高原地球科学卓越中心;中国科学院青藏高原研究所青藏高原环境变化与地表过程重点实验室;中国科学院寒区旱区环境与工程研究所冰冻圈科学国家重点实验室
【刊名】冰川冻土
【关键词】 冰川微生物;温度;盐度;pH
【资助项】国家自然科学基金项目
【ISSN号】1000-0240
【页码】P528-538
【年份】2019
【期号】第2期
【期刊卷】1;|7;|8;|2
【摘要】为了解冰川微生物生长特点,分析了青藏高原木孜塔格冰川、玉珠峰冰川和扎当冰川可培养细菌在不同温度,及木孜塔格冰川可培养细菌在不同盐度和p H下的生长特性.木孜塔格冰川52%的可培养细菌不耐盐,只能在0%盐度下生长,38%的细菌可以在0%~4%/6%盐度培养基中生长,其余细菌可以在0%~1%/2%的盐度培养基中生长,且62%的细菌具有较广的p H值生长范围(p H 5~9);另外,38%的细菌只能在弱酸性(5%)或者只能在弱碱性(33%)培养基中生长.3个冰川可培养细菌生长温度范围均为0~35℃,木孜塔格冰川最适生长温度≤20℃的细菌占其细菌总数的86%,而玉珠峰冰川和扎当冰川最适生长温度≤20℃的细菌则分别占其细菌总数的69%和53%.不同冰川具有不同最适生长温度的细菌的比例不同,同一冰川不同深度相同属类的细菌有相近的生长温度特征、耐盐度和耐酸碱特征.
【全文】 文献传递
青藏高原木孜塔格冰川、玉珠峰冰川及扎当冰川可培养细菌的生理特征
摘 要:为了解冰川微生物生长特点,分析了青藏高原木孜塔格冰川、玉珠峰冰川和扎当冰川可培养细菌在不同温度,及木孜塔格冰川可培养细菌在不同盐度和pH下的生长特性.木孜塔格冰川52%的可培养细菌不耐盐,只能在0%盐度下生长,38%的细菌可以在0%~4%/6%盐度培养基中生长,其余细菌可以在0%~1%/2%的盐度培养基中生长,且62%的细菌具有较广的pH值生长范围(pH 5~9);另外,38%的细菌只能在弱酸性(5%)或者只能在弱碱性(33%)培养基中生长.3个冰川可培养细菌生长温度范围均为0~35℃,木孜塔格冰川最适生长温度≤20℃的细菌占其细菌总数的86%,而玉珠峰冰川和扎当冰川最适生长温度≤20℃的细菌则分别占其细菌总数的69%和53%.不同冰川具有不同最适生长温度的细菌的比例不同,同一冰川不同深度相同属类的细菌有相近的生长温度特征、耐盐度和耐酸碱特征.
关键词:冰川微生物;温度;盐度;pH
doi:10.7522/j.issn.1000-0240.2016.0059
Xing Tingting,Liu Yongqin,Wang Ninglian,et al.The physiological characteristics of culturable bacteria in Muztag、Yuzhufeng and Zadang glaciers on Tibetan Plateau,China[J].Journal of Glaciology and Geocryology,2016,38(2):528-538.[邢婷婷,刘勇勤,王宁练,等.青藏高原木孜塔格冰川、玉珠峰冰川及扎当冰川可培养细菌的生理特征[J].冰川冻土,2016,38(2):528-538.]
0 引言
冰川按年代顺序层层保存了大量微生物,是保存过去微生物的良好载体,也是连接古代与现代环境的纽带[1-2].长期贮存在冰川中的细菌,在自然选择的作用下形成独特的低温适应机制,如产生低温酶、抗冻剂、维持细胞膜稳定性与流动性的不饱和脂肪酸等[3].研究冰川微生物有利于发掘新基因,丰富微生物多样性,揭示过去环境的变迁以及生物进化乃至生命的进化规律,同时也有助于了解微生物对全球气候变化的反馈[4].
我国是中、低纬度地区冰川最发育的国家,其主体是青藏高原冰川[5].青藏高原平均海拔4 000 m以上,其冰川主要集中分布在昆仑山、喜马拉雅山和喀喇昆仑山[6].对青藏高原冰川微生物的研究发现,不同冰川发育不同的微生物种群[7],珠穆朗玛峰北坡海拔6 600~8 000 m表层雪中细菌的优势种群是β-Proteobacteria和Actinobacteria;马兰冰芯细菌优势种群是α-、β-及γ-Proteobacteria、Cytophage-Flexibacter-Bacterioides(CFB);慕士塔格浅冰芯细菌优势种群是α-、γ-Proteobacteria及High G+C gram-positive bacteria(HGC)[8-10]等.此外,不同季节冰川细菌数量和多样性不同,夏季雪中细菌多样性高于冬季,夏季细菌种类多与海洋环境相关,而冬季细菌则具有更强的耐冷性[11-12].这些研究揭示了青藏高原冰川微生物群落多样性、数量分布特征及其与气候环境的关系[9-10,13].但对青藏高原冰川可培养细菌的生理特征的研究较少,对不同冰川可培养细菌的生长温度范围和最适生长温度缺乏对比研究,对其在不同盐度和pH下的生长特点更是少有报道,而这些最基本的生理特征是揭示冰川微生物适应冰川环境及其对气候环境响应机制的基础.
本研究选取青藏高原木孜塔格冰川、玉珠峰冰川及扎当冰川(图1)中的可培养细菌,对比研究了3个冰川共81株可培养细菌在不同温度下的生长特性,及木孜塔格冰川21株可培养细菌在不同盐度和pH下的生长特性,以期初步揭示不同冰川可培养细菌的生理特征.
1 材料和方法
1.1 样品的采集和处理
木孜塔格冰川(36.4°N,87.3°E)位于青藏高原西北部昆仑山主脊,海拔6 973 m,降水量少、气候干燥、辐射强烈,属于大陆型冰川[14-15].玉珠峰冰川(35.5°N,94.2°E)位于高原北部东昆仑地区,海拔6 178 m,气候寒冷干旱[16].扎当冰川(30.5°N,90.6°E)位于念青唐古拉山主峰的东北坡,属于大陆型冰川,冰川区降水主要来自夏季西南季风和局地对流云系[17],最高海拔6 090 m,末端海拔5 515 m.这三个冰川分别位于不同的自然地理带且受不同的大气环流影响,其中,木孜塔格冰川处于高寒荒漠带,主要受西风环流的影响[14],年平均温度-4℃;玉珠峰冰川位于青藏高原北部的高寒草甸带,主要受西风的影响[16],年平均温度-5℃;扎当冰川位于青藏高原中部高寒草甸带,春季主要受西风影响,但夏季受到印度季风的影响更为明显,年平均气温-3.4℃[18].

图1 木孜塔格冰川、玉珠峰冰川以及扎当冰川地理位置
Fig.1 Locations of Muztag Glacier、Yuzhufeng Glacier and Zadang Glacier
玉珠峰冰芯和木孜塔格冰芯分别于2007年和2012年钻取,冰芯样品在中国科学院寒区与旱区环境与工程研究所国家冰冻圈实验室冷库内处理,按5 cm间隔切取冰芯样品,在-20℃超净实验室用灭菌的手术刀切除外层约2 mm的一层,将手术刀在酒精灯上灼烧灭菌后再刮去约2 mm的一层,将切好的冰芯样品放在洁净灭菌的Nalgene瓶中于4℃消融.
雪坑样品的采集是在雪坑剖面每隔10 cm采集微生物雪样,分装入1 L洁净灭菌的Nalgene瓶中,在野外及运输过程中样品一直处于冷冻状态保存.整个过程穿戴洁净无菌服进行所有操作.
1.2 可培养细菌的分离和培养
取冰芯、冰雪融水200 μL涂布于寡营养固体培养基R2A表面,R2A培养基成分为1 L液体培养基中含酵母提取物0.5 g、蛋白胨0.5 g、酸水解酪素0.5 g、葡萄糖0.5 g、可溶性淀粉0.5 g、丙酮酸钠0.3 g、磷酸氢二钾0.3 g、七水硫酸镁0.05 g,调节pH至7.2.R2A固体培养基,是上述1 L培养基中加入15 g琼脂.
4℃培养2~6个月后,挑选平板纯化单菌落,选用通用引物27F和1492R做PCR,进行16S rRNA测序,将得到的所有序列用Blast软件在GenBank数据库进行相似性搜索,获取相近典型菌株的16S rRNA基因序列.同时,将纯化后的单菌落置于液体R2A培养基中,并在200 r·min-1摇床上培养观察,制成原液备用.
1.3 不同温度可培养细菌生理实验
选取木孜塔格冰川、玉珠峰冰川和扎当冰川共81株细菌进行不同温度下的培养(表1).将50 μL细菌原液加入到5 mL R2A液体培养基中,每个样品3个平行样,然后将试管置于不同温度恒温培养箱中培养20 d,振荡均匀测量OD600.不同培养温度设置为0℃、5℃、10℃、15℃、20℃、25℃、30℃和35℃.
1.4 不同盐度可培养细菌生理实验
对木孜塔格冰川21株细菌进行了不同盐度培养(表2).在液体R2A培养基中加入NaCl,使得培养基中盐度值(w/v)分别为1%、2%、3%、4%、5%和6%.将50 μL细菌原液加入到5 mL不同盐度的培养基中,每个样品3个平行样.其中,用于新种鉴定的菌株(B555、B1472、B1555、B1789、B2974)在其最适温度下培养,其余细菌均在室温(18~20℃)下静置培养20 d,振荡均匀测量OD600.
表1 木孜塔格、玉珠峰和扎当冰川可培养细菌生长温度范围和最适生长温度
Table 1 Growth temperature range and the optimum growth temperature of culturable bacteria isolated from Muztag、Yuzhufeng and Zadang Glaciers
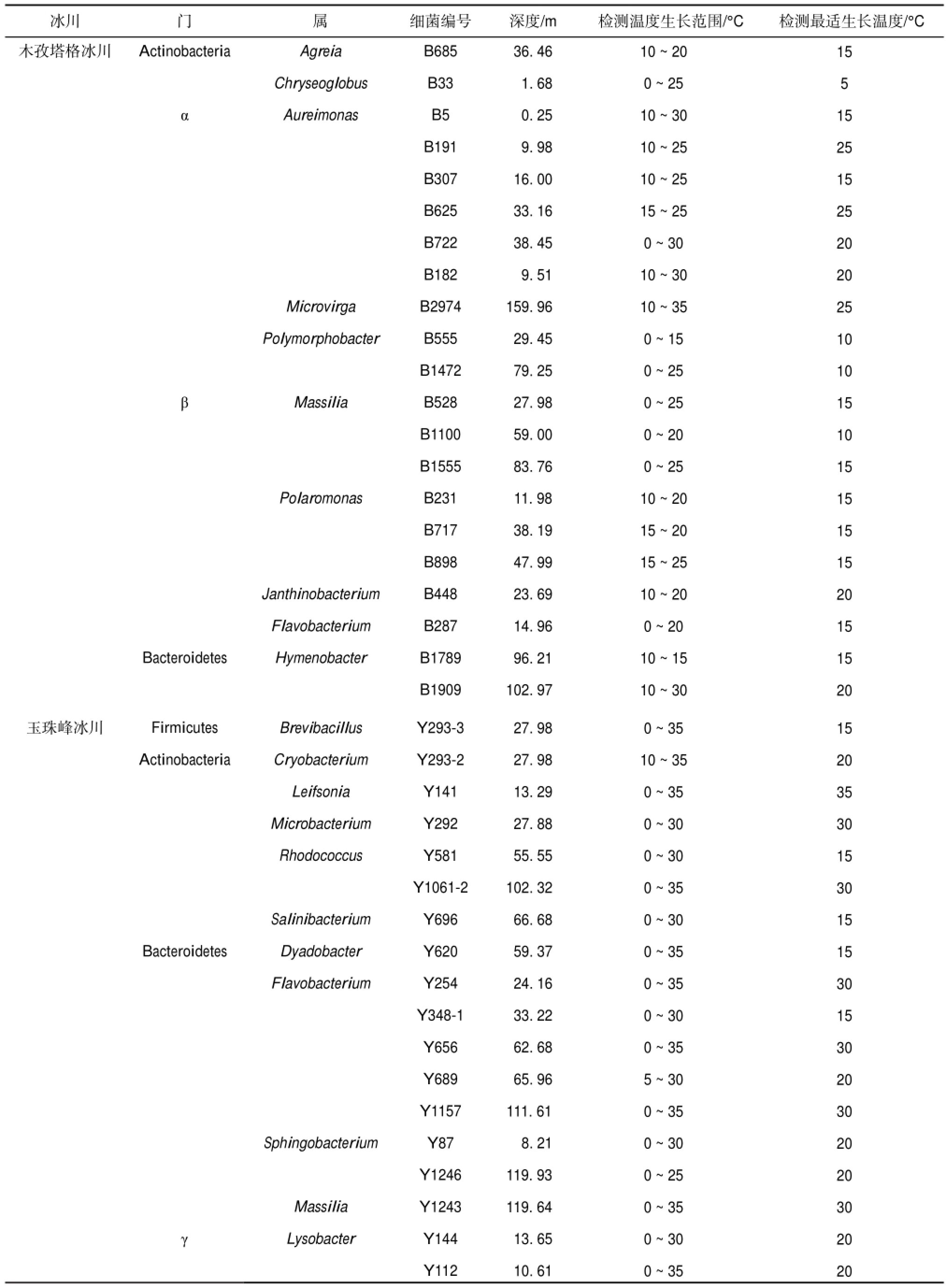

 木孜塔格冰川玉珠峰冰川Actinobacteria α β Bacteroidetes Firmicutes Actinobacteria Bacteroidetes γ Agreia Chryseoglobus Aureimonas Microvirga Polymorphobacter Massilia Polaromonas Janthinobacterium Flavobacterium Hymenobacter Brevibacillus Cryobacterium Leifsonia Microbacterium Rhodococcus Salinibacterium Dyadobacter Flavobacterium Sphingobacterium Massilia Lysobacter B685 B33 B5 B191 B307 B625 B722 B182 B2974 B555 B1472 B528 B1100 B1555 B231 B717 B898 B448 B287 B1789 B1909 Y293-3 Y293-2 Y141 Y292 Y581 Y1061-2 Y696 Y620 Y254 Y348-1 Y656 Y689 Y1157 Y87 Y1246 Y1243 Y144 Y112 36.46 1.68 0.25 9.98 16.00 33.16 38.45 9.51 159.96 29.45 79.25 27.98 59.00 83.76 11.98 38.19 47.99 23.69 14.96 96.21 102.97 27.98 27.98 13.29 27.88 55.55 102.32 66.68 59.37 24.16 33.22 62.68 65.96 111.61 8.21 119.93 119.64 13.65 10.61 10~20 0~25 10~30 10~25 10~25 15~25 0~30 10~30 10~35 0~15 0~25 0~25 0~20 0~25 10~20 15~20 15~25 10~20 0~20 10~15 10~30 0~35 10~35 0~35 0~30 0~30 0~35 0~30 0~35 0~35 0~30 0~35 5~30 0~35 0~30 0~25 0~35 0~30 0~35 15 5 15 25 15 25 20 20 25 10 10 15 10 15 15 15 15 20 15 15 20 15 20 35 30 15 30 15 15 30 15 30 20 30 20 20 30 20 20冰川 门 属 细菌编号 深度/m 检测温度生长范围/°C 检测最适生长温度/°C
木孜塔格冰川玉珠峰冰川Actinobacteria α β Bacteroidetes Firmicutes Actinobacteria Bacteroidetes γ Agreia Chryseoglobus Aureimonas Microvirga Polymorphobacter Massilia Polaromonas Janthinobacterium Flavobacterium Hymenobacter Brevibacillus Cryobacterium Leifsonia Microbacterium Rhodococcus Salinibacterium Dyadobacter Flavobacterium Sphingobacterium Massilia Lysobacter B685 B33 B5 B191 B307 B625 B722 B182 B2974 B555 B1472 B528 B1100 B1555 B231 B717 B898 B448 B287 B1789 B1909 Y293-3 Y293-2 Y141 Y292 Y581 Y1061-2 Y696 Y620 Y254 Y348-1 Y656 Y689 Y1157 Y87 Y1246 Y1243 Y144 Y112 36.46 1.68 0.25 9.98 16.00 33.16 38.45 9.51 159.96 29.45 79.25 27.98 59.00 83.76 11.98 38.19 47.99 23.69 14.96 96.21 102.97 27.98 27.98 13.29 27.88 55.55 102.32 66.68 59.37 24.16 33.22 62.68 65.96 111.61 8.21 119.93 119.64 13.65 10.61 10~20 0~25 10~30 10~25 10~25 15~25 0~30 10~30 10~35 0~15 0~25 0~25 0~20 0~25 10~20 15~20 15~25 10~20 0~20 10~15 10~30 0~35 10~35 0~35 0~30 0~30 0~35 0~30 0~35 0~35 0~30 0~35 5~30 0~35 0~30 0~25 0~35 0~30 0~35 15 5 15 25 15 25 20 20 25 10 10 15 10 15 15 15 15 20 15 15 20 15 20 35 30 15 30 15 15 30 15 30 20 30 20 20 30 20 20冰川 门 属 细菌编号 深度/m 检测温度生长范围/°C 检测最适生长温度/°C
续表1
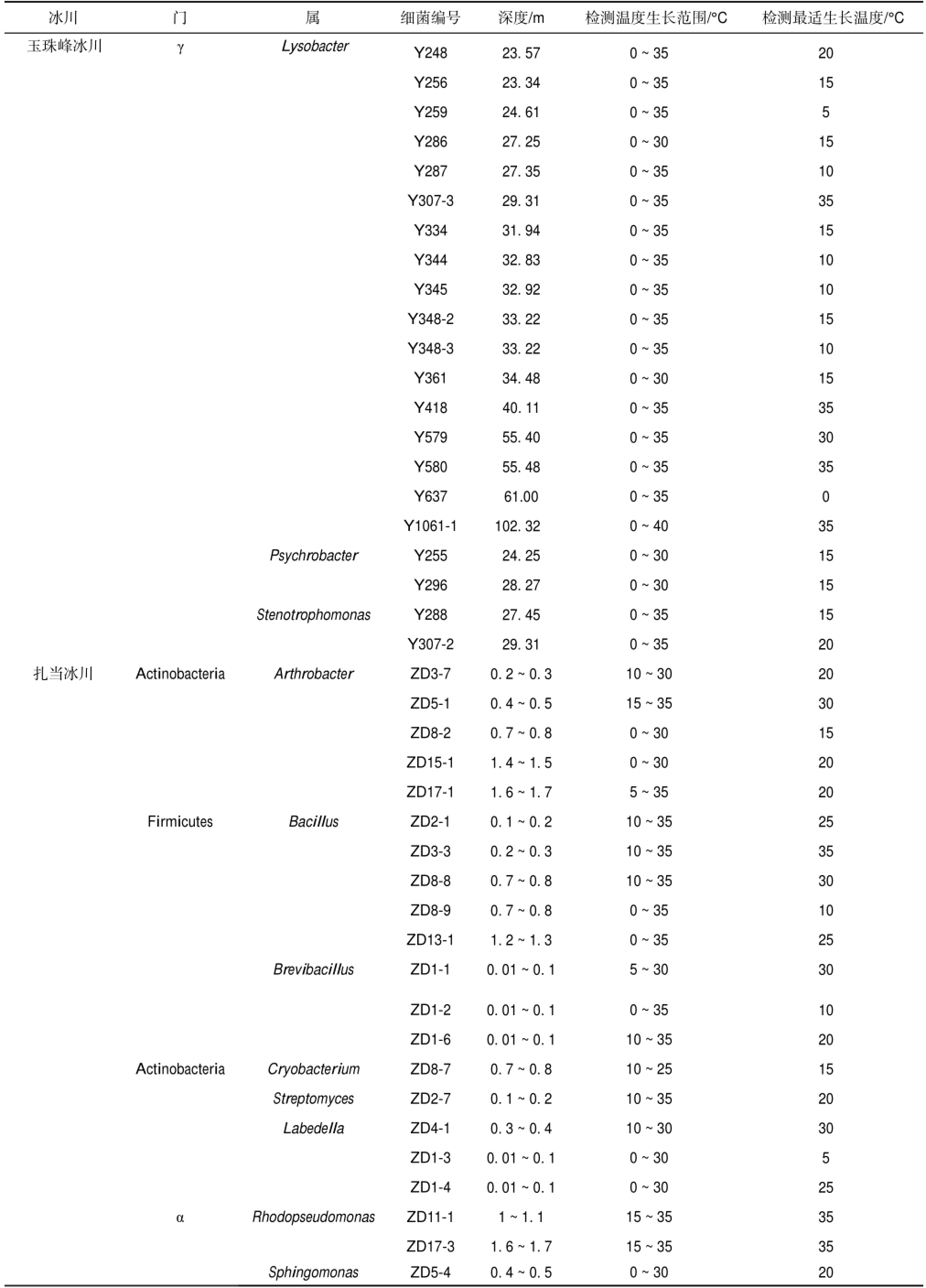
注:α:α-Proteobacteria;β:β-Proteobacteria;γ:γ-Proteobacteria.

 扎当冰川 Actinobacteria Firmicutes Actinobacteria α Psychrobacter Stenotrophomonas Arthrobacter Bacillus Brevibacillus Cryobacterium Streptomyces Labedella Rhodopseudomonas Sphingomonas Y248 Y256 Y259 Y286 Y287 Y307-3 Y334 Y344 Y345 Y348-2 Y348-3 Y361 Y418 Y579 Y580 Y637 Y1061-1 Y255 Y296 Y288 Y307-2 ZD3-7 ZD5-1 ZD8-2 ZD15-1 ZD17-1 ZD2-1 ZD3-3 ZD8-8 ZD8-9 ZD13-1 ZD1-1 ZD1-2 ZD1-6 ZD8-7 ZD2-7 ZD4-1 ZD1-3 ZD1-4 ZD11-1 ZD17-3 ZD5-4 23.57 23.34 24.61 27.25 27.35 29.31 31.94 32.83 32.92 33.22 33.22 34.48 40.11 55.40 55.48 61.00 102.32 24.25 28.27 27.45 29.31 0.2~0.3 0.4~0.5 0.7~0.8 1.4~1.5 1.6~1.7 0.1~0.2 0.2~0.3 0.7~0.8 0.7~0.8 1.2~1.3 0.01~0.1 0.01~0.1 0.01~0.1 0.7~0.8 0.1~0.2 0.3~0.4 0.01~0.1 0.01~0.1 1~1.1 1.6~1.7 0.4~0.5 0~35 0~35 0~35 0~30 0~35 0~35 0~35 0~35 0~35 0~35 0~35 0~30 0~35 0~35 0~35 0~35 0~40 0~30 0~30 0~35 0~35 10~30 15~35 0~30 0~30 5~35 10~35 10~35 10~35 0~35 0~35 5~30 0~35 10~35 10~25 10~35 10~30 0~30 0~30 15~35 15~35 0~30 20 15 5 15 10 35 15 10 10 15 10 15 35 30 35 0 35 15 15 15 20 20 30 15 20 20 25 35 30 10 25 30 10 20 15 20 30 5 25 35 35 20冰川 门 属 细菌编号 深度/m 检测温度生长范围/°C 检测最适生长温度/°C玉珠峰冰川 γ Lysobacter
扎当冰川 Actinobacteria Firmicutes Actinobacteria α Psychrobacter Stenotrophomonas Arthrobacter Bacillus Brevibacillus Cryobacterium Streptomyces Labedella Rhodopseudomonas Sphingomonas Y248 Y256 Y259 Y286 Y287 Y307-3 Y334 Y344 Y345 Y348-2 Y348-3 Y361 Y418 Y579 Y580 Y637 Y1061-1 Y255 Y296 Y288 Y307-2 ZD3-7 ZD5-1 ZD8-2 ZD15-1 ZD17-1 ZD2-1 ZD3-3 ZD8-8 ZD8-9 ZD13-1 ZD1-1 ZD1-2 ZD1-6 ZD8-7 ZD2-7 ZD4-1 ZD1-3 ZD1-4 ZD11-1 ZD17-3 ZD5-4 23.57 23.34 24.61 27.25 27.35 29.31 31.94 32.83 32.92 33.22 33.22 34.48 40.11 55.40 55.48 61.00 102.32 24.25 28.27 27.45 29.31 0.2~0.3 0.4~0.5 0.7~0.8 1.4~1.5 1.6~1.7 0.1~0.2 0.2~0.3 0.7~0.8 0.7~0.8 1.2~1.3 0.01~0.1 0.01~0.1 0.01~0.1 0.7~0.8 0.1~0.2 0.3~0.4 0.01~0.1 0.01~0.1 1~1.1 1.6~1.7 0.4~0.5 0~35 0~35 0~35 0~30 0~35 0~35 0~35 0~35 0~35 0~35 0~35 0~30 0~35 0~35 0~35 0~35 0~40 0~30 0~30 0~35 0~35 10~30 15~35 0~30 0~30 5~35 10~35 10~35 10~35 0~35 0~35 5~30 0~35 10~35 10~25 10~35 10~30 0~30 0~30 15~35 15~35 0~30 20 15 5 15 10 35 15 10 10 15 10 15 35 30 35 0 35 15 15 15 20 20 30 15 20 20 25 35 30 10 25 30 10 20 15 20 30 5 25 35 35 20冰川 门 属 细菌编号 深度/m 检测温度生长范围/°C 检测最适生长温度/°C玉珠峰冰川 γ Lysobacter
1.5 不同pH可培养细菌生理实验
对木孜塔格冰川21株细菌进行了不同pH培养(表3).在液体R2A培养基中加入HCl或者NaOH,使用Sartorius PB-10 pH仪来调节培养基pH,确保培养基的pH分别为5、6、7、8、9、10 和11.pH培养基的配制是先灭菌后调节pH,而后将50 μL细菌原液加入到不同pH值的培养基.其中,用于新种鉴定的菌株(B555、B1472、B1555、B1789、B2974)在其最适温度下培养,其余细菌均在室温(18~20℃)静置培养20 d,振荡均匀测量OD600.
1.6 细菌生长浓度测定
本实验采用光电比浊法测定细菌生长浓度,选取可见光600 nm波段处测定每个试管中培养液的吸光度值.首先,每个试管取200 μL菌液加入到96孔细胞培养板(96 Well Cell Culture Cluster),利用多功能酶标仪(SpectraMax M5 Molecular Devices),选取600 nm波段处进行光电比浊测定OD值.
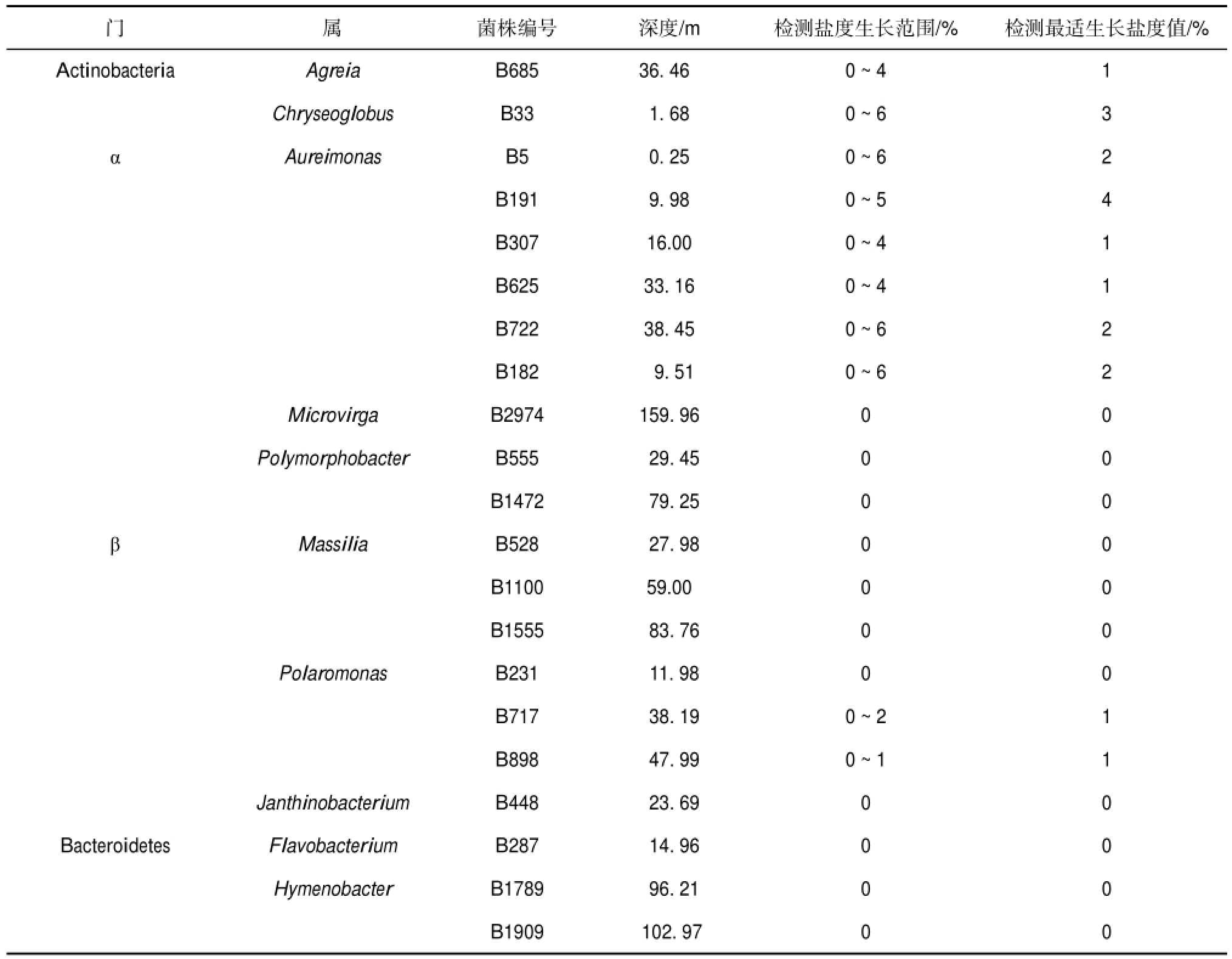
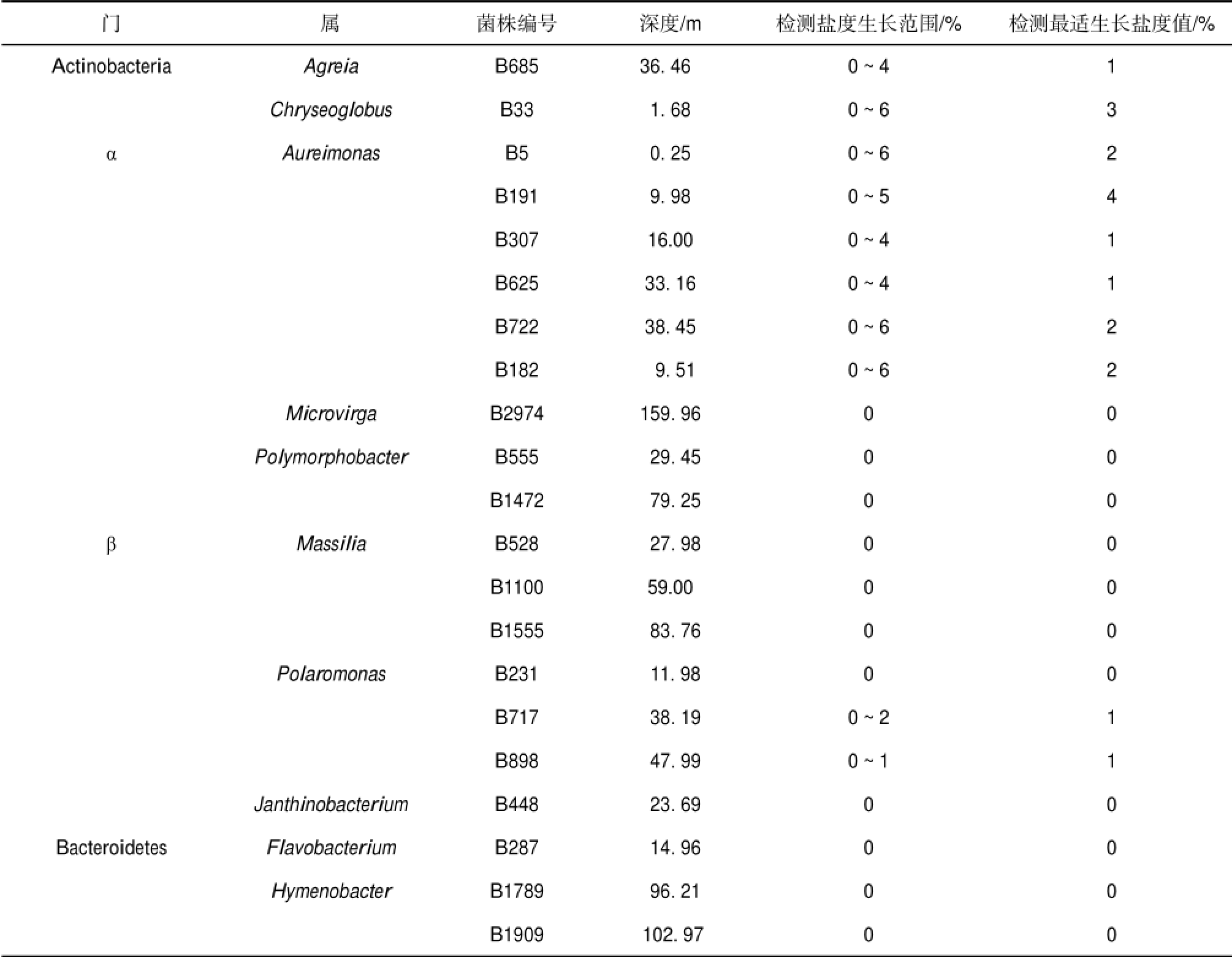
表2 木孜塔格冰川可培养细菌不同盐度生长
Table 2 Growth of culturable bacteria isolated from Muztag glacier in medium with different NaCl
注:α:α-Proteobacteria;β:β-Proteobacteria;γ:γ-Proteobacteria.
 门Actinobacteria α β Bacteroidetes 属Agreia Chryseoglobus Aureimonas Microvirga Polymorphobacter Massilia Polaromonas Janthinobacterium Flavobacterium Hymenobacter菌株编号B685 B33 B5 B191 B307 B625 B722 B182 B2974 B555 B1472 B528 B1100 B1555 B231 B717 B898 B448 B287 B1789 B1909深度/m 36.46 1.68 0.25 9.98 16.00 33.16 38.45 9.51 159.96 29.45 79.25 27.98 59.00 83.76 11.98 38.19 47.99 23.69 14.96 96.21 102.97检测盐度生长范围/% 0~4 0~6 0~6 0~5 0~4 0~4 0~6 0~6 0 0 0 0 0 0 0 0~2 0~1 0 0 0 0检测最适生长盐度值/% 1 3 2 4 1 1 2 2 0 0 0 0 0 0 0 1 1 0 0 0 0
门Actinobacteria α β Bacteroidetes 属Agreia Chryseoglobus Aureimonas Microvirga Polymorphobacter Massilia Polaromonas Janthinobacterium Flavobacterium Hymenobacter菌株编号B685 B33 B5 B191 B307 B625 B722 B182 B2974 B555 B1472 B528 B1100 B1555 B231 B717 B898 B448 B287 B1789 B1909深度/m 36.46 1.68 0.25 9.98 16.00 33.16 38.45 9.51 159.96 29.45 79.25 27.98 59.00 83.76 11.98 38.19 47.99 23.69 14.96 96.21 102.97检测盐度生长范围/% 0~4 0~6 0~6 0~5 0~4 0~4 0~6 0~6 0 0 0 0 0 0 0 0~2 0~1 0 0 0 0检测最适生长盐度值/% 1 3 2 4 1 1 2 2 0 0 0 0 0 0 0 1 1 0 0 0 0
2 分析结果
2.1 冰川可培养细菌在不同温度下的生长
木孜塔格冰川可培养细菌的生长温度范围为0 ~35℃(表1),其中,18株细菌的最适生长温度≤20℃,占86%,最适生长温度最高为25℃(图2a).玉珠峰冰川可培养细菌生长温度范围为0~ 35℃,39株细菌中27株细菌最适生长温度≤20℃,占69%,最适生长温度最高为35℃(图2b).扎当冰川21株可培养细菌的生长温度范围为0~35℃,最适生长温度≤20℃的细菌占52%,最适生长温度最高为35℃(图2c).此外,玉珠峰冰川可以在0℃生长的细菌占其总数的94%,而木孜塔格和扎当冰川在0℃可以生长的细菌数量均占其总数的38%.木孜塔格冰川可培养细菌最低生长温度为0℃、10℃和15℃的细菌分别占38%、48%和14%;玉珠峰冰川可培养细菌最低生长温度为0℃、5℃和10℃的细菌分别占94%、3%和3%;扎当冰川可培养细菌最低生长温度为0℃、5℃、10℃和15℃的细菌分别占38%、10%、38%和14%.
2.2 冰川可培养细菌在不同盐度下的生长
木孜塔格冰川可培养细菌不同盐度培养结果显示(表2),52%的细菌不耐盐,只能在0%的盐度下生长;10%的细菌生长盐度范围分别为0%~1%/ 2%;38%的细菌可以在0%~4%/6%的盐度培养基中生长.Actinobacteria门Agreia属和Chryseoglobus 属2株细菌以及α-Proteobacteria门Aureimonas属的6株细菌具有较高的耐盐度.
2.3 冰川可培养细菌在不同pH下的生长
木孜塔格冰川可培养细菌不同pH培养结果显示(表3),只可以在弱碱性培养基(pH为7~9/10)中生长的细菌占33%;5%的细菌只能在弱酸性培养基(pH为5~7)中生长;62%的细菌具有较广的生长pH范围(pH为5~9/10).相同属类的细菌有着相近的生长pH范围和最适pH,如α-Proteobacteria门Aureimonas属的6株可培养细菌生长pH范围为5~10,而最适生长pH都为5或者6.
表3 木孜塔格冰芯可培养细菌不同pH生长
Table 3 Growth of culturable bacteria isolated Muztag glacier in medium with different pH
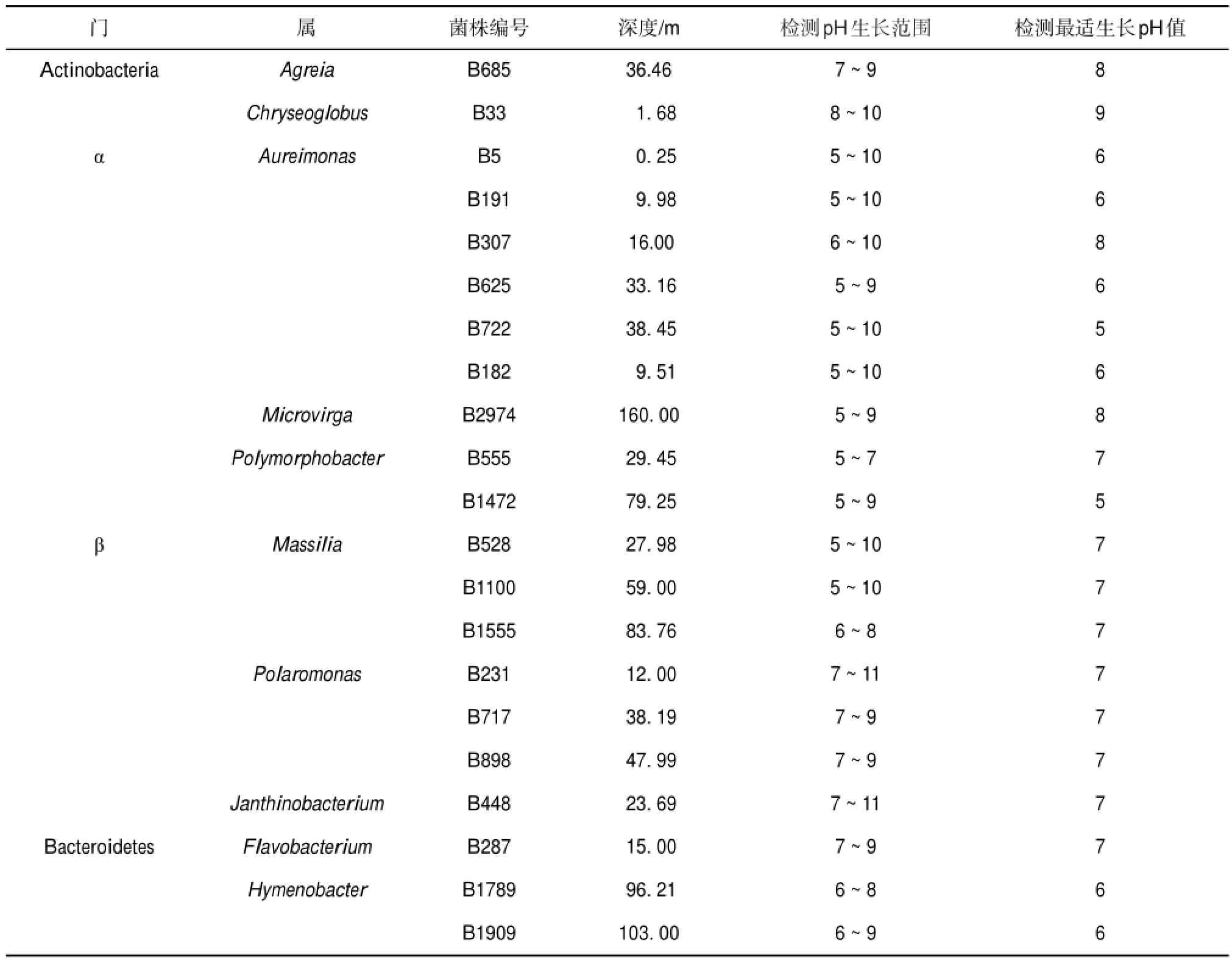
注:α:α-Proteobacteria;β:β-Proteobacteria;γ:γ-Proteobacteria.
门Actinobacteria α β Bacteroidetes 属Agreia Chryseoglobus Aureimonas Microvirga Polymorphobacter Massilia Polaromonas Janthinobacterium Flavobacterium Hymenobacter菌株编号B685 B33 B5 B191 B307 B625 B722 B182 B2974 B555 B1472 B528 B1100 B1555 B231 B717 B898 B448 B287 B1789 B1909深度/m 36.46 1.68 0.25 9.98 16.00 33.16 38.45 9.51 160.00 29.45 79.25 27.98 59.00 83.76 12.00 38.19 47.99 23.69 15.00 96.21 103.00检测pH生长范围7~9 8~10 5~10 5~10 6~10 5~9 5~10 5~10 5~9 5~7 5~9 5~10 5~10 6~8 7~11 7~9 7~9 7~11 7~9 6~8 6~9检测最适生长pH值8 9 6 6 8 6 5 6 8 7 5 7 7 7 7 7 7 7 7 6 6
3 讨论
青藏高原冰川可培养细菌具有较广的温度适应性,生长范围从0~35℃.低温环境中的细菌一般有两类:一类是必须生活在低温环境下且最高生长温度不超过20℃,最适生长温度在15℃,在0℃可以生长繁殖的嗜冷菌;另一类是最高生长温度高于20℃,最适生长温度高于15℃,在0~5℃可生长繁殖的耐冷菌[19].对青藏高原木孜塔格冰川、玉珠峰冰川和扎当冰川可培养细菌生长温度范围和最适生长温度的研究表明,81株实验菌株均为耐冷菌.已有研究证明,青藏高原冰川可培养细菌以耐冷菌为主,如向述荣等[11]对慕士塔格冰芯可培养细菌的研究发现冰芯菌大多数属于耐冷菌和嗜温菌,并且这些嗜温菌可以在-2℃的低温环境中生长;张淑红等[20]采自老虎沟冰川细菌也为耐冷菌.冰川细菌从常温环境被运送并储存在冰川后,细菌会降低最低生长温度和最适生长温度,进化为耐冷菌适应冰川寒冷环境,但这些细菌仍然保留了在常温环境下生长的能力[21].耐冷菌在冰川的低温环境中具有更强的竞争力,冰川微生物是环境选择和生物适应的结果[11].Counnot[22]的研究也证明,冰川低温环境有利于低温菌的生长.木孜塔格冰川中大部分可培养细菌最适生长温度明显低于玉珠峰冰川和扎当冰川(图2),可能是因为木孜塔格冰川海拔最高,且温度最低,较低的冰川温度使得冰川细菌具有更低的最适生长温度,进一步说明不同的冰川环境对细菌的选择作用.此外,冰芯压力高于雪坑,雪坑温度略高于冰芯,随着深度的增加雪坑中Ca2+、Mg2+浓度呈现不同变化趋势[23-24],从雪坑到冰芯,经过低温环境的筛选得到能够适应寒冷环境的细菌.
冰川细菌与分离于冰冻圈外的细菌的最低生长温度和最适生长温度明显不同,如分离于玉珠峰冰川的Lysobacter属(γ-Proteobacter门)的细菌共19株,其温度生长范围为0~35℃,最适生长温度10~20℃居多(68%),最低生长温度为0℃;但分离于土壤和水中的Lysobacter属细菌温度生长范围4~37℃,最适生长温度为20~30℃,最低生长温度4℃[25].另外,分离于木孜塔格冰川的Massilia属(β-Proteobacteria)的3株细菌B528、B1100和 B1555,其温度生长范围分别是0~25℃、0~ 20℃和0~25℃,最适生长温度均是15℃;但分离于空气、土壤、水或者其他环境的Massilia属的细菌的温度生长范围为4~55℃,最适生长温度为28~30℃,最低生长温度为4℃[26-29].与此相似的还有Polaromonas属(β-Proteobacteria)的细菌,分离于常温环境(土壤,自来水等)的细菌的最适生长温度为20℃左右,最低生长温度为4℃;而分离冰雪的Polaromonas属的细菌最适生长温度为15℃,最低生长温度为0℃[30-35].冰川细菌相比较常温环境的细菌具有更低的最适生长温度和最低生长温度.
可培养细菌的最适生长温度并没有随着深度的增加而减小或降低.通过分析冰芯不同深度可培养细菌的生长温度范围和最适生长温度的关系发现,二者并无明显关系并呈现波动分布(图3).玉珠峰冰芯钻孔温度在5~25 m呈现递减趋势[36],但是在该部分分离的细菌的最适生长温度却没有出现相同的趋势.向述荣等[10]对慕士塔格冰芯不同深度可培养细菌温度培养结果也发现,细菌温度生长范围和最适生长温度与深度无明显线性关系.同一冰芯不同深度相同属的细菌具有相近的生长温度范围和最适生长温度,造成细菌生长温度特征差异的主要因素是细菌属类,深度与细菌生长温度范围和最适生长温度无明显关系.这可能是因为不同属类细菌结构存在差异,适应环境能力不同,从而造成生长温度特征不同.
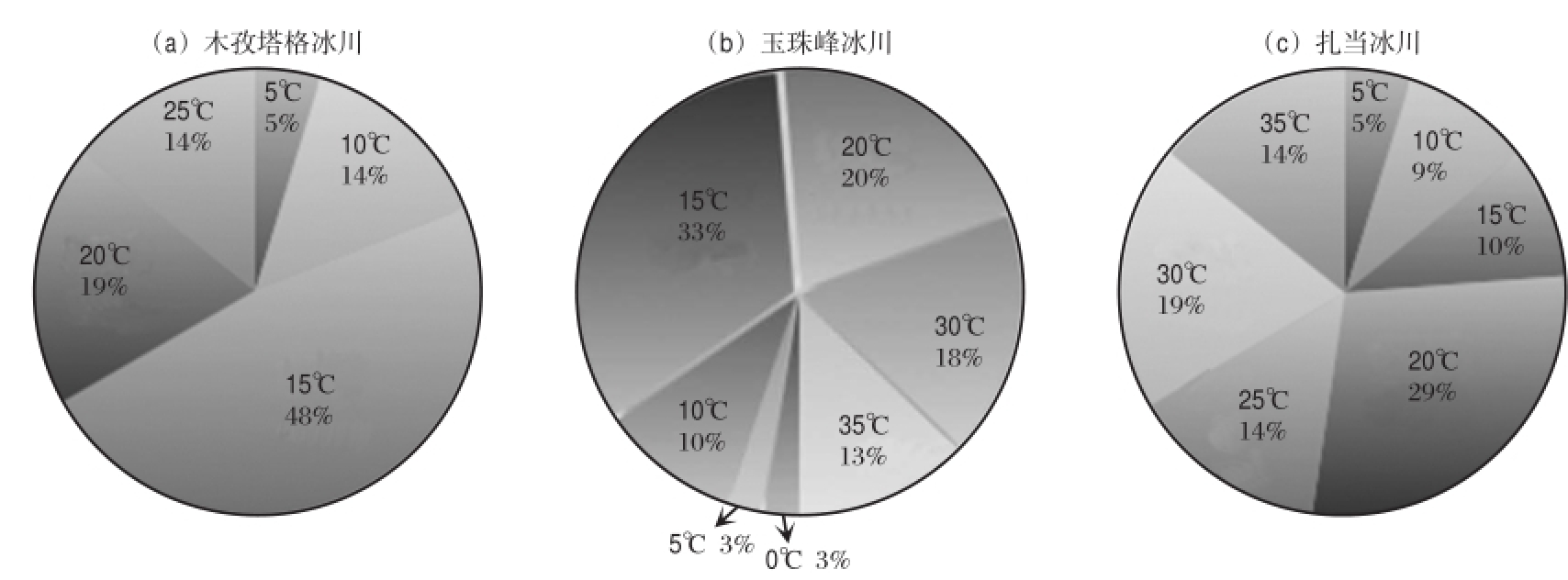
图2 不同冰川具有不同最适生长温度的细菌的比例
Fig.2 The distribution of bacteria with different optimum temperatures in Muztag Glacier (a),Yuzhufeng Glacier(b)and Zadang Glacier(c)
木孜塔格冰川38%的可培养细菌可以在0%~ 4%/6%的广盐度范围内生长(表2),NaCl主要通过影响细胞的渗透压和营养盐的吸收,不同盐度会导致细胞过度伸缩或舒张从而造成细胞死亡,不同细菌对盐度渗透压的适应能力不同[37-38].之前对微生物耐盐度的研究主要集中于海洋细菌,其能分泌耐盐酶,合成、释放无机离子或合成渗压剂(如脯氨酸、甘氨酸、甜菜碱等)来适应海洋高盐度环境[39-40].对青藏高原湖底沉积物中细菌群落的研究表明,盐度与微生物群落分类无显著性关系,随着盐度的升高,可培养细菌的丰度降低[41].与本实验研究一致,在盐度为3%左右时62%冰川可培养细菌不能生长.不同属类的细菌耐盐度存在差异,与不同分类的细菌对盐度渗透压的适应能力不同是一致的[37].
木孜塔格冰川可培养细菌可以在pH 5~9/10的范围内生长的细菌占62%,33%的细菌可以在弱碱性培养基中生长,剩余5%的细菌只能在弱酸性培养基中生长(表3).pH对微生物的影响主要是通过以下两个方面来实现:一是通过影响生物大分子的电荷影响生物活性;二是引起细胞膜电荷变化导致微生物吸收营养物质的能力改变[42].李向应等[43]对青藏高原雪坑和冰芯研究发现雪坑和冰芯呈现弱酸、弱碱性不等;而对天山冰川融水的pH研究得出,从降水到表层雪再到雪坑样品,pH呈现减小的趋势[44].尚天翠等[45]研究发现,慕士塔格细菌在中性或者酸性环境下生长良好,这与慕士塔格冰芯融水弱酸环境(pH=5.2)是一致的.相同属的细菌的耐酸碱范围相似,冰川酸碱度会对细菌的生长产生影响.
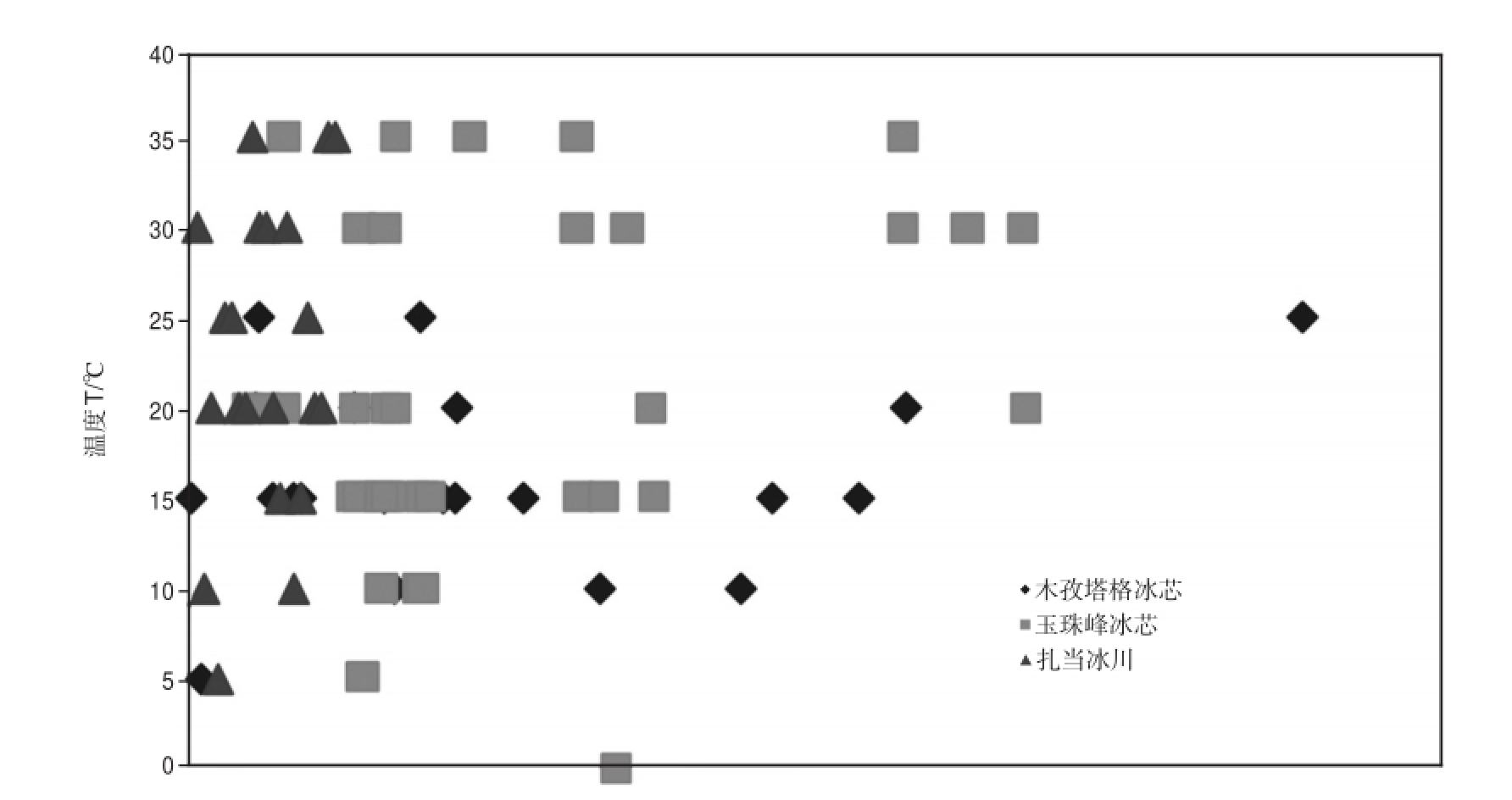
图3 木孜塔格冰芯、玉珠峰冰芯、扎当雪坑不同深度可培养细菌最适生长温度
Fig.3 The optimum growth temperature of culturable bacteria isolated from Muztag Ice Core,Yuzhufeng Ice Core,Zadang Snow pit in different depths
4 结论
青藏高原木孜塔格冰川、玉珠峰冰川及扎当冰川共81株可培养细菌有以下生理特征:木孜塔格、玉珠峰及扎当冰川的可培养细菌都是耐冷菌,均具有较广的生长温度范围;冰川细菌具有较低的最低生长温度和最适生长温度.不同冰川细菌最适生长温度存在差异,同一冰川相同属的细菌温度生长特征相近.木孜塔格冰川可培养细菌耐盐度较低,相同属的细菌耐盐度相似,且木孜塔格冰川可培养细菌一般可以在弱酸、弱碱性培养基中生长.
青藏高原不同冰川可培养细菌具有多样性的生理特征,这些特征一方面说明了冰雪细菌来源于多种环境,另一方面也说明了冰川对细菌的选择作用.青藏高原冰川微生物“储存”在冰雪中,在寒冷环境中仍保持一定的代谢活性,研究其温度、盐度及pH生长特征,为我们后续研究冰雪微生物的代谢途径做好铺垫,也有助于我们更好了解极端环境下微生物的适应机制,进而更好地通过冰川微生物构建还原“古气候”.
参考文献:(References):
[1] Wang Ninglian,Yao Tandong.Contributions of ice core to the past global change research[J].Journal of Glaciology andGeocryology,2003,25(03):275-287.[王宁练,姚檀栋.冰芯对于过去全球变化研究的贡献[J].冰川冻土,2003,25 (03):275-287.]
[2] Zhu Dayun,Wang Jianli.Progress in paleoclimate research on the Tibet Plateau based on ice core records[J].Progress in Geography,2013,32(10):1535-1544.[朱大运,王建力.青藏高原冰芯重建古气候研究进展分析[J].地理科学进展,2013,32(10):1535-1544.]
[3] Lin Xuezheng,Bian Ji,He Peiqing.Molecular mechanism of cold-adaption of polar microorganisms[J].Chinese Journal of Polar Research,2003,15(1):75-82.[林学政,边际,何培青.极地微生物低温适应性分子机制[J].极地研究,2003,15(1):75-82.]
[4] Zhang Shuhong,Hou Shugui,Qin Dahe.Advance in researched of microorganisms in glacial snow and ice on the Qinghai-Tibet Plateau[J].Chinese Journal of Applied and Environmental Biology,2007,13(4):592-596.[张淑红,侯书贵,秦大河.青藏高原冰雪微生物研究进展[J].应用与环境生物学报,2007,13(4):592-596.]
[5] Qin Dahe,Ding Yongjian.Cryopheric changes and their impacts:present,trends and key issues[J].Advances in Climate Change Research,2009,5(4):187-195.[秦大河,丁永建.冰冻圈变化及其影响研究:现状、趋势及关键问题[J].气候变化研究进展,2009,5(4):187-195.]
[6] Liu Zongxiang,Su Zhen,Yao Tandong,et al.Resources and distribution of glaciers on the Tibet Plateau[J].Resource Sciences,2000,22(5):49-52.[刘宗香,苏珍,姚檀栋,等.青藏高原冰川资源及其分布特征[J].资源科学,2000,22(5):49-52.]
[7] Xiang Shurong,Yao Tandong,Chen Yong,et al.Progresses of community distribution of microorganisms in glacier[J].Acta Ecologica Sinica,2006,26(9):3098-3107.[向述荣,姚檀栋,陈勇等.冰川微生物菌群分布的研究概况及其前景[J].生态学报,2006,26(9):3098-3107.]
[8] Liu Yongqin,Yao Tandong,Kang Shichang,et al.Microbial community structure in major habitats above 6000m on Mount Everest[J].Chinese Science Bulletin,2007,52(13):1542-1547.[刘勇勤,姚檀栋,康世昌,等.珠穆朗玛峰北坡6000m以上主要生境细菌群落特征[J].科学通报,2007,52 (13):1542-1547.]
[9] Xiang Shurong,Yao Tandong,An Lizhe,et al.Bacterial flora structure associated with climate change of Malan ice core [J].Chinese Science Bulletin,2004,49(7):1762-1769.[向述荣,姚檀栋,安黎哲,等.马兰冰芯细菌菌群结构变化与气候环境的关系[J].科学通报,2004,49(7):1762-1769.]
[10]Xiang Shurong,Yao Tandong,An Lizhe,et al.16S rRNA sequences and differences in bacteria isolated from the Muztag Ata glacier at increasing depths[J].Sci China Ser D-Earth Sci,2005,35(3):252-262.[向述荣,姚檀栋,安黎哲,等.慕士塔格冰芯可培养细菌的数量分布和主要菌群结构随深度的变化[J].中国科学:D辑地球科学,2005,35(3):252-262.]
[11]Liu Yongqin,Yao Tandong,Kang Shichang,et al.Microbial community structure in moraine lakes and glacial meltwaters,Mount Everest[J].Chinese Science Bulletin, 2006, 51 (11):1287-1296.[刘勇勤,姚檀栋,康世昌,等.珠穆朗玛峰地区东绒布冰川冰雪微生物群落及其季节变化[J].科学通报,2006,51(11):1287-1296.]
[12]Liu Xiaobo,Kang Shichang,Yao Tandong,et al.The Seasonal change of bacterial abundance and diversity in snow of the Guoqu Glacier,Mt.Geladaindong[J].Journal of Glaciology and Geocryology.2009,31(4):634-641.[刘晓波,康世昌,姚檀栋,等.各拉丹冬峰果曲冰川雪中细菌的季节变化特征[J].冰川冻土,2009,31(4):634-641.]
[13]Christner B C,Thompson E M,Thompson L G,et al.Recovery and identification of viable bacteria immured in Glacial Ice [J].Icarus,2000,144:479-485.
[14]Wu Lizong,Li Xin.China glacier information system[M].Beijing:China Ocean Press,2004.[吴立宗,李新,中国冰川信息系统[M].北京:海洋出版社,2004.]
[15]Zhu Meilin,Yao Tandong,Yang Wei,et al.Ice volume and characteristics of sub-glacial topography of the Zhadang Glacier,Nyainqentanglha Range[J].Journal of Glaciology and Geocryology,2014,36(2):268-277.[朱美林,姚檀栋,杨威,等.念青唐古拉山扎当冰川冰储量估算及冰下地形特征分析[J].冰川冻土,2014,36(2):268-277.]
[16]Li Quanlian,Wang Ninglian,Wu Xiaobo,et al.Environmental records within the snowpits in the Yuzhufeng Glacier and Xiao Dongkemadi Glacier on the Tibetan Plateau[J].Journal of Glaciology and Geocryology.2011,33(1):47-54.[李全莲,王宁练,武小波,等.青藏高原玉珠峰冰川和小冬克玛底冰河攒雪坑中的环境记录[J].冰川冻土,2011,33(1):47-54.]
[17]Yu Guangming,Xu Jianzhong,Kang Shichang,et al.Mineral and elemental characteristics of microparticles with different size fraction in snow pits from Zadang Glacier[J].Environmental Science,2011,32(11):3264-3270.[余光明,徐建中,康世昌,等.扎当冰川雪坑中不同粒径微粒元素和矿物组成特征[J].环境科学,2011,32(11):3264-3270.]
[18]You Qinglong,Kang Shichang,Tian Keming,et al.Preliminary analysis on climatic features at Mt.Nyainqentanglha,Tibetan Plateau[J].Journal of Mountain Science,2007,25 (4):497-504.[游庆龙,康世昌,田克明,等.青藏高原念青唐古拉峰地区气候特征初步分析[J].山地学报,2007,25(4):497-504.]
[19]Xin Xiuming,Ma Yanhe.Psychrophilies and psychrotrophs [J].Microbiology China,1999,26(2):155-156.[辛秀明,马延和.嗜冷菌和耐冷菌[J],微生物学通报,1999,26 (2):155-156.]
[20]Zhang Shuhong,Yang Hongchao,Liu Xiuhua,et al.Preliminary study on enzymatic.activities of four glacier snow bacteria [J].Biotechnology,2011,21(2):80-85.[张淑红,杨红超,刘秀花等.四株冰川雪细菌产酶能力的初步研究[J].生物技术,2011,21(2):80-85.]
[21]Shen Liang,Yao Tandong,Liu Yongqin,et al.Downwardshifting temperature range for the growth of snow-bacteria on glaciers of the Tibetan Plateau[J].Geomicrobiology Journal,2014,31(9):779-787.
[22]Gounot A M.Effects of temperature on the growth of psychrophilic bacteria from glaciers[J].Can J Microbiol,1976,22 (6):839-46.
[23]Liu Yaping,Hou Shugui,Ren Jiawen,et al.Distribution features of borehole temperature in the Miaoergou Flat Topped Glacier,East Tianshan Mountains[J].Journal of Glaciology and Geocryology,2006,28(5):668-671.[刘亚平,侯书贵,任贾文,等.东天山庙儿沟平顶冰川钻孔温度分布特征[J].冰川冻土,2006,28(5):668-671.]
[24]Yu Guangming,Xu Jianzhong,Kang Shichang,et al.Postdepositional features of the insoluble microparticles on the Laohugou Glacier No.12,Qilian Mountains[J].Journal of Glaciology and Geocryology,2014,36(6):1394-1402.[余光明,徐建中,康世昌,等.祁连山老虎沟12号冰川微粒在沉积后过程中的变化特征[J].冰川冻土,2014,36(6):1394-1402.]
[25]Christensen P,Cook F D.Lysobacter,a new genus of nonfruitin,gliding bacteria with a high base ratio[J].International Journal of Systematic Bacteriology,1978,28(3),367-393.
[26]Shen Liang,Liu Yongqin,Wang Ninglian,et al.Massilia yuzhufengensisi sp.nov.,isolated from an ice core[J].International Journal of System and Evolutionary Microbiology,2013,63,1285-1290.
[27]Gallego V,Sanchez-Porro C,GarcıaM T,et al.Massilia aurea sp.nov.,isolated from drinking water[J].International Journal of System and Evolutionary Microbiology,2006,56,2449-2453.
[28]Weon H Y,Hong S B,Jeon YA,et al.Massilia niabensis sp.nov.and Massilia niastensis sp.nov.,isolated from air samples [J].International Journal of System and Evolutionary Microbiology,2006,59,1656-1660.
[29]Zul D,Wanner G,Overmann J.Massilia brevitalea sp.nov.,a novel betaproteobacterium isolated from lysimeter soil[J].International Journal of System and Evolutionary Microbiology,2008,58,1245-1251.
[30]Irgens R L,Gosink J J,Staley J T.Polaromonas vacuolata gen.nov.,sp.nov.,a psychrophilic,marine,gas vacuolate bacterium from Antarctica[J].International Journal of System and Evolutionary Microbiology,1996,46,822-826.
[31]Margesin R,Sproer C,Zhang D C,et al.Polaromonas glacialis sp.nov.and Polaromonas cryoconiti sp.nov.,isolated from alpine glacier cryoconite[J].International Journal of System and Evolutionary Microbiology,2012,62,2662-2668.
[32]Jeon C O,Park W,Ghiorse W C,et al.Polaromona naphthalenivorans sp.nov.,a naphthalene-degrading bacterium from naphthalene contaminated sediment[J].International Journal of System and Evolutionary Microbiology,2014,54,93-97.
[33]Maria S,Nicolai P.Polaromonas hydrogenivorans sp.nov.,a psychrotolerant hydrogen-oxidizing bacterium from Alaskan soil [J].International Journal of System and Evolutionary Microbiology,2007,57,616-619.
[34]Kämpfer P,Busse H,Falsen E.Polaromonas aquatica sp.nov.,isolated from tapwater[J].International Journal of System and Evolutionary Microbiology,2006,56,605-608.
[35]Weon H Y,Yoo S H,Hong S B,et al.Polaromonas jejuensis sp.nov.,isolated from soil in Korea[J].International Journal of System and Evolutionary Microbiology,2008,58,1525-1528.
[36]Wang Ninglian,Xu Baiqing,Pu Jianchen,et al.Discovery of the water-rich ice layers in glaciers on the Tibetan Plateau and its environmental significances[J].Journal of Glaciology and Geocryology,2013,35(6):1371-1381.[王宁练,徐柏青,蒲健辰,等.青藏高原冰川内部富含水冰层的发现及其环境意义[J].冰川冻土,2013,35(6):1371-1381.]
[37]Zhou Tao,Xu Xiangrong,Xu Hao,et al.Research of bacterial biofilm growth based on osmotic pressure theory[J].Journal of Jinggangshan University(Natural Sciences),2014,35 (5):33-37.[周涛,徐向荣,徐浩,等.基于渗透压理论对细菌生物膜生长的研究[J].井冈山大学学报(自然科学版),2014,35(5):33-37.]
[38]Dong Haitao,Sun Hongyi.Effect of NaCl stress on ecophysiological characteristics of transplanting seeding of Stevia rebaudiana Bertoni[J].Journal of Glaciology and Geocryology,2015,37(2):538-544.[董海涛,孙宏义.NaCl胁迫对甜叶菊移栽苗生理生态特性的影响[J].冰川冻土,2015,37 (2):538-544.]
[39]Luo Zhuhua,Pei Yaowen,Huang Xiangling,et al.Physiological and ecological characteristics of the deep sea microorganisms in tropical Pacific Ocean[J].Acta Oceanologica Sinica.2006,28(5):130-137.[骆祝华,裴耀文,黄翔玲,等.热带太平洋深海微生物的若干生理生态特征[J].海洋学报(中文版),2006,28(5):130-137.]
[40]Yu Yong,Li Huiron,Chen Bo,et al.Advance of research for sea ice bacteria[J].Chinese Journal of Polar Research,2005,17(1):75-85.[俞勇,李会荣,陈波,等.海冰细菌研究进展[J].极地研究,2005,17(1):75-85.]
[41]Xiong Jinbo,Liu Yongqin,Lin Xiangui,et al.Geographic distance and pH drive bacterial distribution in alkaline lake sediments across Tibetan Plateau[J].Environmental Microbiology,2012,14(9):2457-2466.
[42]Chen Yanfei.pH to uygur biology influence[J].Journal of Taiyuan Normal University(Natural Science Edition),2009,8 (3):121-124.[陈燕飞.pH对微生物的影响[J].太原师范学院学报(自然科学版),2009,8(3):121-124.]
[43]Li Xiangying,Qin Dahe,Han Tianding,et al.Progress in precipitation chemistry in cryophere regions of Western China [J].Progress in Geography.2011,30(1):3-16.[李向应,秦大河,韩添丁,等.中国西部冰冻全地区大气降水化学的研究进展[J].地理科学进展,2011,30(1):3-16.]
[44]Zhou Ping,Zhang Mingjun,Li Zhongqin,et al.Seasonal variations of pH and electrical conductivity in precipitation and snow on the Glaciers of Tianshan Mountains,China[J].Arid Land Geography,2010,4(33):518-524.[周平,张明军,李忠勤,等.中国天山冰川区降水、积雪pH和电导率季节变化特征分析[J].干旱区地理,2010,4(33):518-524.]
[45]Shang Tiancui,Xu Bingliang,Xiang Shurong,et al.Physicological characteristics of isolates from Muztag Ata ice core and sequence analysis of Staphylococcus equorum Muzt-D84 cspA gene[J].Journal of Gansu Agricultural University,2007,42 (4):65-70.[尚天翠,徐秉良,向述荣,等.慕士塔格冰芯细菌的生理特征以及葡萄球菌Muzt-D84 csp A基因序列分析[J].甘肃农业大学学报,2007,42(4):65-70.]
The physiological characteristics of culturable bacteria in Muztag、Yuzhufeng and Zadang Glaciers on Tibetan Plateau,China
Abstract:In order to reveal the characteristics of glacier-microorganism,we assayed the culturable bacteria isolated from Muztag Glacier,Yuzhufeng Glacier and Zadang Glacier at different temperatures.Besides,culturable bacteria isolated from Muztag Glacier was observed at different salinity and pH.Salinity endurance of 52%of Muztag Glacier culturable bacteria was 0%,and 38%bacteria could grow in the medium of NaCl density with 0%~4%/6%,while the other bacterial salinity tolerance range was 0%~1%/2%.Muztag Glacier culturable bacteria had wide range pH tolerance(5~9)occurring 62%,while bacteria only grew in weak acid(5%)or weak alkaline(33%)medium accounting for 38%.Growth-temperature range of bacteria in these three glaciers was 0 ~35℃.The Muztag bacterial optimum temperature which was below 20℃(including 20℃)accounted for 86%,while Yuzhufeng Glacier and Zadang Glacier were 69%and 53%,respectively.The percent of different optimum temperatures in different glaciers were different.Culturable bacteria belonged to one genus isolated from same glacier had similar growth-temperature range,salinity and pH tolerance in different depths.
Key words:glacier microorganism;temperature;NaCl;pH
中图分类号:Q935
文献标志码:A
文章编号:1000-0240(2016)02-0528-11
收稿日期:2015-12-28;
修订日期:2016-03-08
基金项目:国家自然科学基金项目(41425004;41190084;41371084)资助