珠江三角洲东部晚第四纪微体动物群的古环境意义*
【类型】期刊
【作者】吴洁,刘春莲,张素青,杨婷婷,殷鉴(中山大学地球科学系;广东省地质过程与矿产资源探查重点实验室)
【作者单位】中山大学地球科学系;广东省地质过程与矿产资源探查重点实验室
【刊名】中山大学学报(自然科学版)
【关键词】 珠江三角洲;有孔虫;介形类;晚第四纪;古环境
【资助项】国家自然科学基金资助项目(40872024);德国学术交流中心(daad)与国家留学基金委(csc)国际合作交流资助项目
【ISSN号】0529-6579
【页码】P45-52
【年份】2019
【期号】第6期
【期刊卷】1;|7;|8;|4;|5;|2
【摘要】珠江三角洲是世界上主要河口三角洲之一,自晚更新世开始形成,其沉积物中保存了丰富的微体动物壳体,它们是重建晚第四纪古环境极为有效的指标。本文对珠江三角洲东部PRD17孔的微体动物群(有孔虫和介形类)进行了分析,共发现有孔虫14属29种,介形类14属17种,将其分别划分为3种生态类型。根据微体动物群的组成特征,并结合测年数据、岩性、沉积物粒度及颜色反射率等特征,重建了PRD17孔晚第四纪以来的环境演化。约43 400 cal a B.P.以前,钻孔所在地发育河流相沉积。晚更新世玉木亚间冰期开始,南海海平面回升,本区开始形成第四纪以来的第一个海相层位。从微体动物群的组成和丰度可识别出3次小规模的海平面波动。约35 400~30 900 cal a B.P.为晚更新世最大海侵时期,微体动物群的丰度和分异度都达到峰值。约25 900 cal a B.P.以后,海水逐渐退出。末次冰盛期的海平面大幅下降,导致本区上更新统沉积物暴露地表遭受风化剥蚀,形成珠江三角洲普遍发育的花斑状黏土层。由于钻孔所在地地势相对较高,受全新世海侵的影响较珠江三角洲其它地区晚,直到6 000 cal a B.P.左右才重新接受沉积,初期发育上潮间带沉积。约3 700 cal a B.P.开始,逐渐变为受潮水影响较显著的潮坪环境。
【全文】 文献传递
珠江三角洲东部晚第四纪微体动物群的古环境意义*
摘 要:珠江三角洲是世界上主要河口三角洲之一,自晚更新世开始形成,其沉积物中保存了丰富的微体动物壳体,它们是重建晚第四纪古环境极为有效的指标。本文对珠江三角洲东部PRD17孔的微体动物群(有孔虫和介形类)进行了分析,共发现有孔虫14属29种,介形类14属17种,将其分别划分为3种生态类型。根据微体动物群的组成特征,并结合测年数据、岩性、沉积物粒度及颜色反射率等特征,重建了PRD17孔晚第四纪以来的环境演化。约43 400 cal a B.P.以前,钻孔所在地发育河流相沉积。晚更新世玉木亚间冰期开始,南海海平面回升,本区开始形成第四纪以来的第一个海相层位。从微体动物群的组成和丰度可识别出3次小规模的海平面波动。约35 400~30 900 cal a B.P.为晚更新世最大海侵时期,微体动物群的丰度和分异度都达到峰值。约25 900 cal a B.P.以后,海水逐渐退出。末次冰盛期的海平面大幅下降,导致本区上更新统沉积物暴露地表遭受风化剥蚀,形成珠江三角洲普遍发育的花斑状黏土层。由于钻孔所在地地势相对较高,受全新世海侵的影响较珠江三角洲其它地区晚,直到6 000 cal a B.P.左右才重新接受沉积,初期发育上潮间带沉积。约3 700 cal a B.P.开始,逐渐变为受潮水影响较显著的潮坪环境。
关键词:珠江三角洲;有孔虫;介形类;晚第四纪;古环境
河口三角洲地区处于海陆过渡地带,对气候与环境变化非常敏感,尤其是反映海平面变化及其所导致的一系列环境变化的沉积记录保存良好,为研究晚第四纪环境变化提供了极为有利的条件。有孔虫和介形类作为河口三角洲环境中两类重要的生物类群,它们的化石和现代类型数量均很多,且生态适应范围也很广。因此,尤为适合作为河口三角洲环境分析的指标[1-2]。自20世纪80年代起,部分学者已陆续开始利用这两类微体生物重建珠江三角洲地区晚第四纪古环境[3-8],基本认识了微体动物群对于海平面变化等古环境演变的生态响应,但在沉积相和沉积旋回的划分上还存在一些分歧。这主要是由于珠江三角洲晚第四纪不同断块的构造运动具有明显的地区差异性[9-10],不同构造单元的沉积物中记录的海侵旋回及环境变迁存在着较大的时空差异。本文通过分析珠江三角洲广州—番禺断块的PRD17孔中有孔虫和介形类组合特征,并结合测年数据、岩性、沉积物粒度及颜色反射率等特征,重建了PRD17孔晚更新世以来的环境演化。经与前人工作对比,探讨了海平面变化和地壳升降对海侵旋回的综合影响,为进一步了解珠江三角洲晚第四纪环境演化提供依据。
1 采样与方法
PRD17孔位于珠江三角洲广州番禺鱼窝头镇(22°51′07″N,113°26′30″E),孔深33.34 m。在孔深26.43~1.14 m的岩芯中共取116个样品用于微体化石分析,取样间距约为20 cm。顶部和底部岩芯未取样,因前者可能受人工扰动,后者主要为砾石及基岩风化层。沉积物样品在60 ℃下烘干,称取约100 g干样在烧杯中用水浸泡~24 h,待样品散开后分别用0.063和0.125 mm孔径筛过筛。其中大于0.125 mm的沉积物是含有孔虫和介形类的主体,将用于微体化石的鉴定和统计(以100 g干样计算)。
沉积物粒度是判别沉积环境尤其是水动力条件的良好指标[11]。本文对PRD17孔深27.46~1.10 m的岩芯进行了粒度分析取样,共计120个样品。每个样品取10 g左右,放入60 ℃烘箱烘干后称取干样质量。随后加入约10 mL φ=10%的HCl溶液,以除去样品中的碳酸盐,充分搅拌直至样品中无气泡产生。接着加入约10 mL体积分数φ为20%的H2O2溶液,以去除有机质,搅拌使其充分反应。静置12 h后抽去上层清液,加入10 mL的0.1 mol/L的(NaPO3)6作为分散剂,搅拌后静置12 h抽去上层清液,最后用水稀释、清洗样品,保留沉积物悬浊液待上机测试。本次粒度测试采用英国产Mastersizer 2000激光粒度仪,其测量范围为0.02~2 000 μm。对于粒径大于2 000 μm的样品,先过2 mm湿筛,然后称量筛上物的干样质量,细粒部分仍用激光粒度仪进行测试,两部分数据合并得出完整的粒度分布。
沉积物颜色是沉积物最醒目的标志,它主要取决于沉积物的矿物组分和有机质含量,也可反映沉积物形成时的氧化还原程度[12]。本文采用香港美能达公司生产的CR-400/410手持色差仪对PRD17孔岩芯进行了高分辨率的颜色反射率测定,测点间距为2 cm,测试范围为孔深33.24~1.12 m的岩芯。获得了两个颜色反射率指数L*和a*的变化曲线,分别代表样品亮度和红绿彩度。前者可反映沉积物的碳酸盐和有机质含量的变化[13]。L*越高,表明碳酸盐含量越高,有机质含量越低,反之亦然。后者可反映不同价态铁的氢氧化物含量及碳酸镁含量。较高的a*值代表岩心颜色较红,反映较强的氧化沉积环境,而a*值较低则代表沉积环境氧化性较弱,还原性较强[14]。
沉积物年龄采用常规14C测年法,利用超低本底液体闪烁仪(1220 Quantulus)对富有机质的沉积物样品进行测定,在中国科学院广州地球化学研究所进行测试。共获得7个14C年龄值,采用CALIB软件校正为日历年龄(图1),其它各深度的年龄根据等间距插值法估算。文中讨论皆使用校正后年龄。
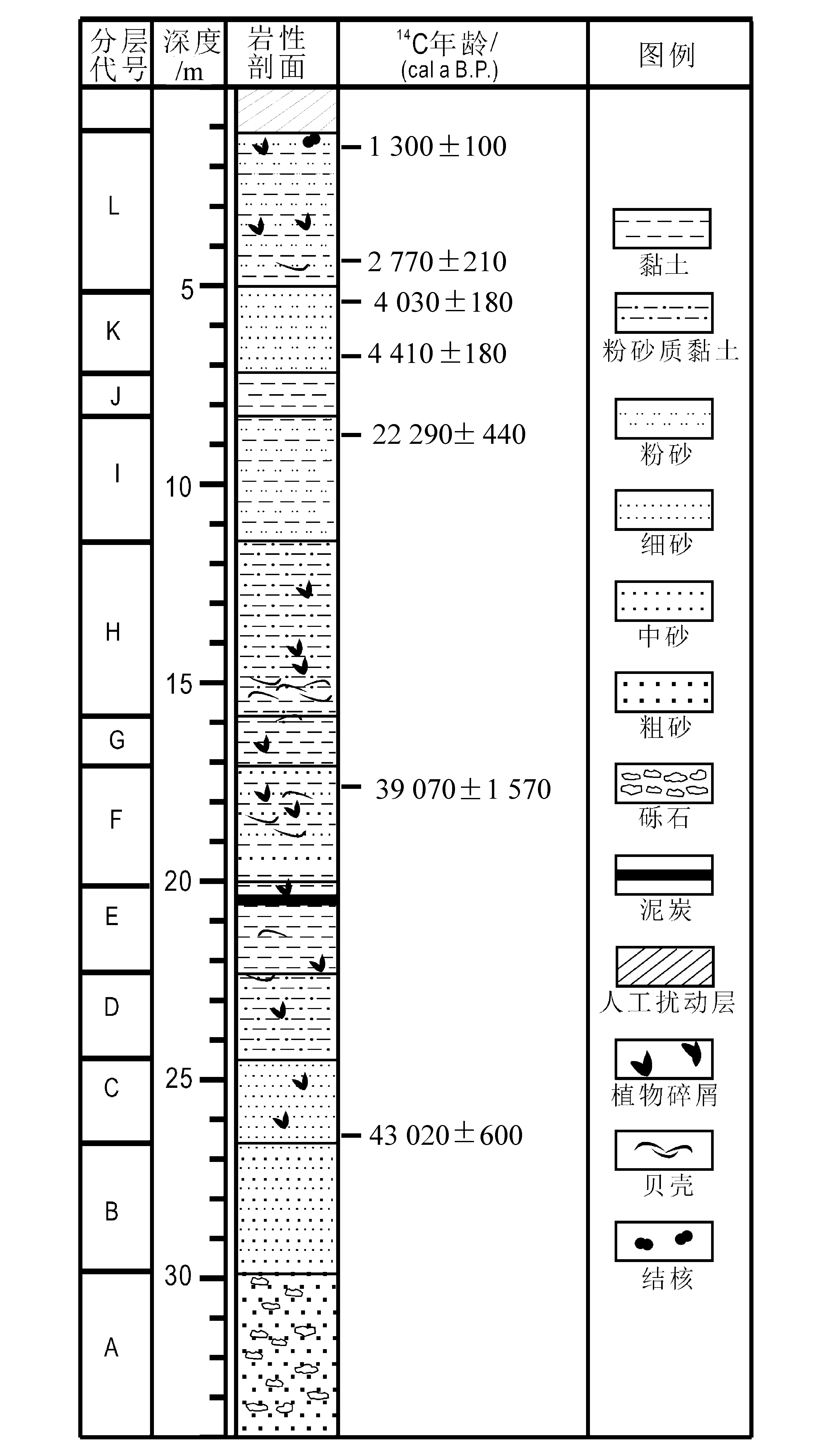
图1 PRD17孔岩性特征及14C测年结果
Fig.1 Lithological characteristics and calibrated dates of borehole PRD17
2 岩性特征
将岩芯从中间剖开,详细观察岩性、沉积构造及其它组分(包括软体动物壳体及其碎片、炭屑和结核等)特征,结合粒度分析结果,将全孔岩芯从下至上划分为12层(图1)。
A.孔深33.27~29.82 m:灰黄色中粗砂,含砾石。平均粒径分布范围为0.99~1.34Φ,分选性差。
B.孔深29.82~26.63 m:灰白色中细砂。平均粒径范围为3.31~4.53Φ,分选性中等。上部岩性特征及颜色反射率值发生明显变化。
C.孔深26.63~24.48 m:深灰色细砂。沉积物粒度波动较大,平均粒径范围为1.92~4.60Φ,分选性中等。24.57~24.79和25.95~26.55 m两个层位含有较多腐木碎屑。
D.孔深24.48~22.34 m:深灰色粉砂质黏土。平均粒径范围为3.21~5.43Φ,分选性较差。22.40 m处发现大块腐木(2 cm×3 cm)。上部含少量贝壳碎片。孔深23.30 m处开始持续出现微体动物化石。表明海平面上升,钻孔所在地开始受到海水影响。
E.孔深22.34~20.13 m:深灰色黏土。平均粒径范围为3.93~5.00Φ,分选性差。上部含一层厚约16 cm的泥炭层。一些层位含贝壳。微体化石总丰度逐渐增加。
F.孔深20.13~17.05 m:深灰色黏土质中细砂。平均粒径范围为1.48~4.44Φ,分选性较差。含大量贝壳碎片。微体动物群丰度和分异度先降后升。
G.孔深17.05~15.91 m:深灰色黏土。平均粒径范围为3.63~4.92Φ,分选性差。部分层位含贝壳碎片。微体化石数量短暂下降。
H.孔深15.91~11.85 m:深灰色粉砂质黏土。平均粒径范围为3.98~4.84Φ,分选性较差。底部含较多炭屑。微体化石丰度和分异度均达到峰值。
I.孔深11.85~8.42 m:深灰色黏土与粉砂互层。平均粒径范围为4.23~5.69Φ,分选性较差。微体化石数量逐渐下降,直至为零。表明海水逐渐退出本区。
J.孔深8.42~7.49 m:花斑状黏土,多呈黄褐色。平均粒径范围为2.28~5.16Φ,分选性较差。此阶段对应末次冰盛期的陆相风化剥蚀环境。
K.孔深7.49~5.14 m:深灰色黏土质粉砂。平均粒径范围为2.99~5.04Φ,分选性较差。未见任何微体化石、贝壳碎片及植物碎屑。
L.孔深5.14~1.05 m:深灰色粉砂与黏土互层。平均粒径范围为3.04~5.26Φ,分选性中等。部分层位含有贝壳碎片和腐木碎屑。有孔虫和介形类仅零星出现, 且多为随潮水搬运而来的异地分子。孔深1.05 m以上为人工扰动层,不做分析讨论。
3 微体动物群分析
PRD17孔共发现有孔虫14属29种,介形类14属17种。两类微体动物群的丰度分布特征基本相似。钻孔底部较粗粒沉积物及其上部的沼泽沉积中未发现微体化石。自孔深23.30 m处开始同时出现有孔虫和介形类壳体。17.05~13.98 m有孔虫化石较丰富,介形类主要集中于23.30~15.06 m。孔深11.07 m以上微体化石仅零星出现(图2)。

图2 PRD17孔沉积特征、微体古生物分布及古环境演化阶段划分
Fig.2 Depositional characteristics, microfaunal distribution and environmental stages of borehole PRD17
将微体动物群按生态特征划分为不同的类型,通过讨论不同生态类型动物群的垂直分布特征可有效重建钻孔所在地的古环境[15-17]。本文也采取这种方法,将钻孔中出现的有孔虫和介形类按照各属种的生态特征划分为3种类型,通过讨论3种不同生态类型的动物群的垂直分布来重建PRD17孔晚更新世以来的环境变化。
3.1 有孔虫
Group1:浅海型。是一类常见于大陆架浅海海域的有孔虫。本钻孔中仅出现Massilina penglaiensis和Globigerina bulloides两个浅海种,且丰度较低。
Group2:海相-半咸水型。多分布于中盐-多盐条件的水域,水体盐度通常大于25‰,水深范围主要为20~50 m。本孔中主要有以下12种属于该类型, Cavarotalia annectens、Elphidium advenum、Elphidium hispidulum、Cribrononion asiaticum、Quinqueloculina seminulangulata、Pararotalia nipponica、Rosalina bradyi、Elphidium magellanicum、Quinqueloculina sp.、Florilus decorus、Nonion boueanum、Elphidium sp.。
Group3:滨岸广盐型。水深分布范围0~50 m,以20 m以内的浅水水域为主。这类属种可适应的盐度范围很广,低盐至正常盐度条件下均可生存。包括Ammonia beccarii、Ammonia tepida、Ammonia koeboeensis、Ammonia sp.、Ammonia convexidorsa、Quinqueloculina akneriana rotunda、Ammonia pauciloculata、Ammonia sobrina、Ammonia takanabensis、Ammonia ketienziensis、Ammonia maruhasii、Ammonia confertitesta Zheng、Cellanthus craticulatus、Rectobolivina raphana、Poroeponides incrassatus,其中Ammonia beccarii、Ammonia tepida两个种的数量约占PRD17孔有孔虫总丰度的40%,在现代南海和东海沿岸半咸水环境中,这两类也是主要的优势种[18-19]。
3.2 介形类
PRD17孔介形类属种划分的3个生态类型与有孔虫属种的基本一致。
Group1:浅海型。包括Stigmatocythere rosemani、Macrocypris decora、Acitinocythereis kisarazuensis、Abyssocythere regalis,本孔中丰度较低。
Group2:海相-半咸水型。主要有Bicorncythere bisanensis、Alocopocythere kendengensis、Pistocythereis bradyformis、Sinocythere sinensis、Keijella hodgii、Neosinocythere elongata、Neomonoceratina dongtaiensis、Nipponocythere obesa、Alocopocythere profusa、Cornucoquimba tosaensis。其中 Pistocythereis bradyformis常见于多盐至真盐环境[19],要求的水深也大于其它种[20]。
Group3:滨岸广盐型。包括Sinocytheridea impressa、Bicorncythere leizhouensis及 Neomonoceratina iniqua三个种。
4 古环境重建
根据上述岩性和微体动物群分布特征,并结合测年数据、沉积粒度及沉积物颜色反射率(L*,a*)等数据,PRD17孔晚更新世以来的环境演化可分为5个阶段。
4.1 约43 400 cal a B.P. 以前(33.27~26.63 m)
未发现有孔虫和介形类化石,亦未含任何生物贝壳碎片及腐木碎屑。底部以中粗砂为主,含砾石。推测可能为水动力条件较强的河道沉积。向上沉积物粒度逐渐减小为中细砂,表明水动力条件有所减弱。这与王建华等[21]得出的GZ-2孔(PRD17孔东南侧)晚更新世海侵之前为河床相沉积的结论较一致。
4.2 约43 400~21 900 cal a B.P.(26.63~8.42 m)
晚更新世玉木亚间冰期,海平面逐渐回升。海侵初期(约43 400~41 700 cal a B.P.),本区形成深灰色的沼泽相沉积。约41 700 cal a B.P.,海水开始全面进入本区,钻孔所在地持续发育河口湾环境,直至约21 900 cal a B.P.。期间共发生3次小规模的海平面波动,导致水体条件发生变化,进而影响了微体动物群的组成和丰度。晚更新世中期最大海侵时期发生于约35 400~30 900 cal a B.P.。
4.2.1 约43 400~40 300 cal a B.P.(26.63~20.13 m) 孔深23.30 m处首次出现微体化石。有孔虫丰度和分异度均较低。主要属种为滨岸广盐型分子Ammonia beccarii和Ammonia tepida,其丰度均不高于100枚/100 g。介形类丰度较低,但分异度较高。主要属种为滨岸广盐型分子Sinocytheridea impressa和Bicorncythere leizhouensis,其丰度均不高于300枚/100 g。伴随出现丰度较低的海相-半咸水型分子Bicorncythere bisanensis、Alocopocythere kendengensis、Pistocythereis bradyformis、Neomonoceratina dongtaiensis。其余属种仅少量出现。此阶段出现了介形类海相型分子,但数量均较低(< 25枚/100 g),可能为异地搬运分子。
开始于距今5万年的晚更新世玉木亚间冰期,南海海平面开始上升[22],但钻孔所在地可能处于古河流间的低洼处,海平面上升导致排水不畅而形成富含有机质的沼泽相沉积。直到约41 700 cal a B.P.(孔深23.30 m)时海水经古珠江水道进入本区[23],钻孔所在地开始发育河口湾沉积。随着水深进一步加大,尽管微体动物化石丰度及分异度较低,但数量逐渐开始增多,且出现了海相种和海相-半咸水种。据此推断珠江三角洲晚更新世中期海侵可能在约41 700 cal a B.P.已经开始,这与李平日等[24]认为本区晚更新世中期海进开始年限比38,400 a B.P.还早的结论一致。
4.2.2 约40 300~39 400 cal a B.P.(20.13~18.20 m) 有孔虫和介形类丰度均有所下降。有孔虫以滨岸广盐型分子Ammonia beccarii为主,丰度一般低于100枚/100 g。介形类广盐型分子Sinocytheridea impressa和Bicorncythere leizhouensis的数量大大下降,它们的丰度均小于100枚/100 g。伴随出现的介形类海相-半咸水型分子Bicorncythere bisanensis、Alocopocythere kendengensis和Pistocythereis bradyformis大都仅30~80枚/100 g。其余属种仅零星出现。前一时期出现的介形类海相型分子消失。
此阶段为海平面短暂下降期,持续时间约为1.0 ka,前期出现的介形类海相种已难觅踪迹,仅有少数以Ammonia beccarii为代表的广盐性分子,说明水深和盐度较前一阶段明显下降。
4.2.3 约39 400~38 000 cal a B.P.(18.20~17.05 m) 微体化石丰度和分异度均快速增加。有孔虫除滨岸广盐型分子Ammonia beccarii和Ammonia tepida外,还包括了海相-半咸水型分子Cavarotalia annectens、Elphidium advenum、Elphidium hispidulum,丰度一般为100~400枚/100 g。有孔虫海相型分子首次出现。介形类广盐型分子Sinocytheridea impressa和Bicorncythere leizhouensis的数量显著提高,它们的丰度大都高于200枚/100 g。介形类海相-半咸水型分子Bicorncythere bisanensis、Alocopocythere kendengensis和Pistocythereis bradyformis也都高于100枚/100 g。介形类浅海型分子Stigmatocythere rosemanis仅零星出现。
海平面重新快速上升,有孔虫和介形类的分异度呈现显著上升趋势,且此阶段有孔虫和介形类的海相分子数量都达到较高水平。说明尽管本次海侵持续时间较前一阶段(约43 400~40 300 cal a B.P.)短,但规模更大,受海水影响更显著,有利于海相-半咸水属种的繁荣。
4.2.4 约38~000~35 400 cal a B.P.(17.05~15.91 m) 微体动物群化石经历了一个短暂的丰度和分异度下降。未发现有孔虫海相型分子。滨岸广盐型分子和海相-半咸水型分子的数量也较低,大都低于200枚/100 g。介形类仅零星出现广盐型分子Sinocytheridea impressa、Bicorncythere leizhouensis和少量海相-半咸水型分子Neomonoceratina dongtaiensis、Alocopocythere kendengensis、Pistocythereis bradyformi。它们的数量均低于50枚/100 g。
Pistocythereis bradyformis等要求水深较大、盐度偏高的海相-半咸水型分子明显减少,反映规模较小、时间较短的海退过程,水体盐度和深度均有所下降。
4.2.5 约35 400~30 900 cal a B.P.(15.91~ 13.98 m) 微体化石的丰度和分异度再次大幅增加,并在孔深15.06 m处达到峰值。尽管有孔虫和介形类主要优势种仍为广盐型分子,但海相-半咸水型分子的数量也较其它阶段高。有孔虫广盐性分子主要包括Ammonia beccarii、Ammonia tepida、Ammonia koeboeensis、Ammonia sp.、Ammonia convexidorsa、Quinqueloculina aknerian arotunda、Ammonia pauciloculata、Ammonia takanabensis、Ammonia ketienziensis、Ammonia maruhasii、Ammonia confertitesta Zheng、Cellanthus craticulatus、Rectobolivina raphana和Poroeponides incrassatus。其中Ammonia beccarii、Ammonia tepida、Ammonia koeboeensis三个种的平均丰度均超过400枚/100 g。海相-半咸水型分子主要有Cavarotalia annectens、Elphidium advenum、Elphidium hispidulum、Cribrononion asiaticum、Elphidium magellanicum、Quinqueloculina sp.、Florilus decorus。其中Cavarotalia annectens和Elphidium advenum的丰度多高于200枚/100 g。介形类广盐性分子以Sinocytheridea impressa、Bicorncythere leizhouensis和Neomonoceratina iniqua为主,它们的丰度都不小于200枚/100 g。伴随出现的介形类海相-半咸水型分子主要有Neomonoceratina dongtaiensis、Alocopocythere kendengensis、Pistocythereis bradyformi。
海平面再次上升,有孔虫和介形类的总丰度和分异度都大大增加,达到整个钻孔的峰值。尽管海相分子数量较少,但此阶段半咸水-广盐性分子的空前繁荣仍然可代表珠江三角洲晚更新世中期最大海侵时期。这与前人的研究结果较为一致[21, 24-26]。
4.2.6 约30 900~21 900 cal a B.P.(13.98~8.42m) 有孔虫丰度和分异度开始显著下降。海相型分子不再出现。广盐性分子和海相-半咸水型分子的丰度都大大低于前一阶段。广盐性分子最大壳体数一般不超过300枚/100 g。海相-半咸水型分子数不高于50枚/100 g。孔深11.85 m处开始有孔虫极少出现。本阶段未发现介形类化石。
有孔虫和介形类的丰度及分异度开始持续下降,直至约25 900 cal a B.P.(11.85 m)。此后研究钻孔中晚更新世地层再无微体化石出现,标志着玉木亚间冰期基本结束。
关于晚更新世海侵层的年代现在仍无定论,有学者认为本次海侵层对应MIS3期[17, 27-29]。Zong等[30]则认为由于此海相层在末次冰期时暴露地表遭受风化,故所测得的年龄比其真实年龄普遍偏年轻,且MIS3期的最高海平面低于现代海平面,因此本阶段应属于MIS5时期沉积的海侵层。然而,也有其它证据表明在MIS3时期的海平面可能比现代海平面更高[31-36]。如果PRD17孔的测年数据可信,本次海侵层应属于MIS3期。珠江三角洲其它钻孔也记录了本区晚更新世曾发生过较为广泛的海侵[21, 37]。本次晚更新世海侵的高海平时期约为35 400~30 900 cal a B.P.,与中国东海及长江三角洲晚更新世高海平面期的年代基本一致[29, 38],说明此次海侵具有普遍性。
4.3 约21 900~6 000 cal a B.P.(8.42~7.49 m)
黄褐色花斑状黏土层,颜色反射率再一次处于高值区,指示较强的氧化环境。从测年数据上看,花斑状黏土层形成于末次冰盛期的大海退时期并延续至早全新世。随着末次冰盛期的到来,全球海平面发生大幅下降,珠江三角洲乃至整个南海北部陆架均处于陆相风化剥蚀环境[17, 35],上更新统沉积物在冰盛期暴露地表从而形成风化层[7, 39]。根据本孔花斑黏土层上、下伏沉积物的沉积速率推算出其形成年龄约为21 900~6 000 cal a B.P.。这与黄镇国等[35]提出的珠江口晚第四纪钻孔中普遍发育的两个风化层的上风化层时间(MIS 2)较一致。
末次冰期结束后,全球气候变暖,海平面开始回升,海平面上升初期,海水尚未侵入珠江三角洲地区。随着海平面进一步上升,海水开始入侵,但由于珠江三角洲晚第四纪存在多个构造断块,且各个断块的垂直差异运动强度不尽相同[10],导致不同断块接受海侵沉积的时间不同[7]。庄畅等[8]采自北江与东江河口交汇处的PRD16孔约7 500 cal a B.P.开始接受全新世海侵沉积。而珠江三角洲南部平原大鳌沙地区的PRD05孔[17]全新世初期约10 100 cal a B.P.就已经开始接受海侵沉积。本文的研究孔PRD17位于广州—番禺断块,在晚更新世和全新世的抬升速率明显高于其他断块,地势相对较高,开始受海水影响的时间较晚,根据花斑黏土层上伏沉积物的沉积速率,推测出PRD17孔全新统底部沉积物年龄大致为6 000 cal a B.P.。
4.4 约6 000~ 3 700 cal a B.P.(7.49~5.14 m)
沉积物以黏土质粉砂为主,未发现微体化石记录,推测为上潮间带环境。
4.5 3 700~1 100 cal a B.P. (5.14~ m)
随着全新世后期广州—番禺断块进一步下沉[10],钻孔所在地受潮水影响渐为显著,发育潮坪环境。沉积特征表现为深灰色粉砂与黏土互层,部分层位偶见零星有孔虫和介形类壳体,应属随潮水搬运而来的异地分子。
从PRD17孔微体动物群记录看,本孔晚更新世海侵层较全新世更为发育,这与邻近区域内其它钻孔(PRD16孔,GZ-2孔)全新世海侵层更为发育的状况不甚相同[8, 21]。根据珠江三角洲晚第四纪第一期砂砾层(约40.0~33.0 ka B.P.)的分布特征所绘制的该时期古河谷位置图[22,40-41]可知,此时,狮子洋水道尚未形成,古东江与古北江相汇于PRD17孔所处的沙湾附近。本区地势较低洼的河谷广泛地接受了晚更新世海侵沉积(约43 400~22 000 cal a B.P.)。晚更新世后期随着萝岗-太平断裂上盘下降[40],狮子洋水道开始形成,东江不再经由沙湾而直接汇入狮子洋,逐渐偏离PRD17孔所在地,加之晚更新世海侵的结束,PRD17孔的水深进一步下降。此外,PRD17孔所处的广州—番禺断块自全新世以来一直处于抬升,其抬升速率绝对值之和超过2 mm/a[10]。故尽管全新世海侵最高海平面较晚更新世更高,但对PRD17孔的影响仍未达到晚更新世的程度。
5 结 论
对珠江三角洲PRD17孔沉积物和微体动物群的分析表明,受海平面变化和河流作用的共同影响,研究区晚更新世以来经历了多次环境变化。距今4万多年以前,钻孔所在地发育河流相沉积。玉木亚间冰期全球性海侵,导致河口位置逐渐向内陆推进,本区开始发育沼泽相沉积,直至约41 700 cal a B.P.,海水开始进入本区,钻孔所在地持续发育河口海湾环境,41 700~25 900 cal a B.P.为晚更新世大海侵时期,期间共发生3次小规模的海平面波动,导致水体条件发生变化,从而影响了微体动物群的组成和丰度。更新世中期最大海侵时期发生于35 400~30 900 cal a B.P.,PRD17孔微体动物群的丰度和分异度都达到最大值。25 900 cal a B.P.以后,海水开始逐渐退出本区。随着末次冰盛期的到来,海平面大幅下降,本区上更新统沉积物开始暴露地表遭受风化剥蚀。末次冰期结束后,海平面回升,PRD17孔所在的广州—番禺断块由于地势相对较高,开始受海侵影响的时间较珠江三角洲其它地区晚,6 000 cal a B.P.左右才开始重新接受沉积,初期发育上潮间带环境,约3 700 cal a B.P.以来,部分层位偶见零星微体化石,本区可能逐渐变为受潮水影响较为显著的潮坪沉积。
参考文献:
[1]BOOMER I, HORNE D J, SLIPPER I J. The use of ostracodes in paleoenvironmental studies, or what can you do with an ostracode shell? [J]. Paleontological Society Papers, 2003, 9: 153-179.
[2]MENDES I, GONZALEZ R, DIAS J M A, et al. Factors influencing recent benthic foraminifera distribution on the Guadiana shelf (Southwestern Iberia) [J]. Marine Micropaleontology, 2004, 51 (1): 171-192.
[3]袁家义,陈木宏,余家桢,等. 珠江三角洲北部晚第四纪更新世以来的沉积特征[J]. 海洋通报, 1988, 7 (2): 49-61.
[4]赵焕庭. 珠江河口演变[M]. 北京:海洋出版社, 1990.
[5]YIM W W S, HE X X. Distribution of foraminifera in late quaternary sediments off High Island, Hong Kong [J]. Journal of Southeast Asian Earth Sciences, 1991, 6 (1): 1-11.
[6]陈木宏,赵焕庭,温孝胜,等. 伶仃洋 L2 和 L16 孔第四纪有孔虫群与孢粉化石带特征及其地质意义[J]. 海洋地质与第四纪地质, 1994, 14 (1): 11-22.
[7]刘春莲,杨婷婷,吴洁,等. 珠江三角洲晚第四纪风化层稀土元素地球化学特征[J]. 古地理学报, 2012, 14 (1): 125-132.
[8]庄畅,刘春莲,吴洁,等. 珠江三角洲PRD16孔微体生物记录与晚更新世以来的古环境重建[J]. 中山大学学报:自然科学版, 2013, 52 (3): 128-32.
[9]钟建强,詹文欢,古森昌,等. 珠江三角洲新构造运动与地壳稳定性分析[J]. 华南地震,1996,16 (12): 57-63.
[10]陈伟光,赵红梅,常郁,等. 珠江三角洲晚第四纪垂直构造运动速率[J]. 地震地质, 2001, 23 (4): 581-587.
[11]朱筱敏.沉积岩石学[M]. 北京:石油工业出版社,2008.
[12]吴艳宏,李世杰. 湖泊沉积物色度在短尺度古气候研究中的应用[J]. 地球科学进展, 2004, 19 (5): 789-792.
[13]刘升发, 王昆山,刘焱光,等. 长江口泥质区沉积物颜色反射率指示的近百年来东亚季风变迁[J]. 沉积学报,2013, 31 (2): 331-339.
[14]黄维, 翦知湣, BÜHRING C. 南海北部ODP1144站颜色反射率揭示的千年尺度气候波动[J]. 海洋地质与第四纪地质, 2004, 23 (3): 5-10.
[15]MAZZINI I, ANADON P, BARBIERI M, et al. Late Quaternary sea level changes along the Tyrrhenian coast near Orbetello (Tuscany, central Italy): palaeoenvironmental reconstruction using ostracods [J]. Marine Micropaleontology, 1997, 37: 289-311.
[16]TA T K O, NGUYEN V L, TATEISHI M, et al. Sedimentary facies, diatom and foraminifer assemblages in a late Pleistocene-Holocene incised valley sequence from the Mekong River Delta, Bentre Province, Southern Vietnam: the BT2 core [J]. Journal of Asian Earth Sciences, 2001, 20: 83-94.
[17]LIU C L, FÜRSICH F T, WU J, et al. Late Quaternary paleoenvironmental changes documented by macrofauna and shell stable isotopes in the southern Pearl River Delta plain, South China [J]. Journal of Palaeogeography, 2013, 2 (4): 344-361.
[18]李淑鸾. 珠江口底质中有孔虫埋葬群的分布规律[J]. 海洋地质与第四纪地质,1985, 5 (2): 83-104.
[19]汪品先,章纪军,赵泉鸿,等. 东海底质中的有孔虫和介形虫[M]. 北京:海洋出版社,1988.
[20]刘春莲,FÜSICH F T,董艺辛,等. 珠江三角洲PRD05孔的高分辨率介形类记录与晚第四纪古环境重建[J].古地理学报,2008, 10 ( 3) : 313-322.
[21]王建华,曹玲珑,王晓静,等. 珠江三角洲万顷沙地区晚第四纪沉积相与古环境演变[J]. 海洋地质与第四纪地质,2009, 29 (6): 35-41.
[22]黄镇国,李平日,张仲英,等. 珠江三角洲地区晚更新世以来海平面变化及构造运动问题[J]. 热带地理,1982, 2 (1) : 29-38.
[23]陈国能,张珂,贺细坤,等. 珠江三角洲晚更新世以来的沉积—古地理[J]. 第四纪研究,1994(1): 67-74.
[24]李平日,黄镇国,张仲英,等. 广东东部晚更新世以来的海平面变化[J]. 海洋学报:中文版, 1987, 9 (2): 216-222.
[25]耿秀山. 中国东部晚更新世以来的海水进退[J]. 海洋学报, 1981, 3 (1): 114-130.
[26]汪品先,闵秋宝,卞云华,等. 我国东部第四纪海侵地层的初步研究[J]. 地质学报, 1981, 55 (1): 1 -13.
[27]赵焕庭,陈木宏,余家桢,等. 珠江三角洲海进层微体古生物的初步研究[J]. 热带海洋,1987,6(1): 28-36.
[28]陈荣华. 浙江沿海地区第四纪海侵 [J]. 海洋学报,1992, 14(3): 76-85.
[29]王张华,赵宝成,陈静,等. 长江三角洲地区晚第四纪年代地层框架及两次海侵问题的初步探讨[J]. 古地理学报,2008, 10(1): 99-110.
[30]ZONG Y, YIM W W S, YU F, et al. Late Quaternary environmental changes in the Pearl River mouth region, China [J]. Quaternary International, 2009, 206 (1): 35-45.
[31]刘尚仁. 从古海平面遗迹看晚更新世中国海的高海面高度 [J]. 热带海洋, 1997, 16(3):23 -31.
[32]BELLUOMINI G, CALDARA M, CASINI C, et al. The age of Late Pleistocene shorelines and tectonic activity of Taranto area, Southern Italy [J].Quaternary Science Re views, 2002, 21: 525-547.
[33]杨达源,陈可锋,舒肖明. 深海氧同位素第3阶段晚期长江三角洲古环境初步研究 [J]. 第四纪研究,2004, 24(5): 525-530.
[34]HANEBUTH T J J, SAITO Y, TANABE S, et al. Sea levels during late marine isotope stage 3 (or older?) reported from the Red River Delta (northern Vietnam) and adjacent regions [J]. Quaternary International, 2006, 145/146: 119-134.
[35]黄镇国,蔡福祥. 珠江口晚第四纪埋藏风化层及其环境意义[J]. 第四纪研究,2007, 27 (5): 828-833.
[36]GRACIA F J, RODRíGUEZVIDAL J, CáCERES L M, et al. Diapiric uplift of an MIS 3 marine deposit in SW Spain: Implications for Late Pleistocene sea level reconstruction and palaeogeography of the Strait of Gibraltar [J]. Quaternary Science Reviews, 2008, 27: 2219-2231.
[37]郑卓, 王建华. 珠江三角洲北部晚第四纪孢粉植物群的古环境意义[J]. 热带海洋, 1998,17(3): 1-9.
[38]张军强, 唐璐璐, 邹昊. 晚更新世以来古气候与海平面变化在东海地区的响应[J]. 海洋湖沼通报, 2008 (1): 25-31.
[39]蓝先洪. 珠江三角洲晚第四纪沉积特征[J]. 沉积学报, 1996, 14 (2): 155-161.
[40]CHEN G, ZHANG K, LI L, et al. Development of the Pearl River Delta in SE china and its relations to reactivation of basement faults [J]. Journal of Geosciences of China, 2002, 14 (1): 17-24.
[41]姚衍桃,詹文欢,刘再峰,等. 珠江三角洲的新构造运动及其与三角洲演化的关系[J]. 华南地震,2008,28(1): 29-41.
Palaeoenvironmental Significance of Late Quaternary Microfaunas in the Eastern Pearl River Delta
Abstract:The Pearl River Delta, formed since the Late Pleistocene, is one of the major delta in the world. There are abundant microfaunas preserved in sediments of the Pearl River Delta, which have been proved to be essential for reconstructing Quaternary environment. In this study, Microfaunas (foraminifers and ostracods) of borehole PRD17 from the eastern Pearl River Delta have been analyzed quantitatively. A total of 29 foraminiferal species belonging to 14 genera and 17 ostracod species referred to 14 genera were identified. They can be divided into three different groups with respect to their ecological ranges. Based on the comprehensive analysis of microfaunas, lithological and sediment grainsize characteristics of core samples, palaeoenvironmental changes since the Late Pleistocene can be reconstructed. Before ~43 400 cal a B.P., a fluvial environment was recorded in the borehole until the late Wurm Interglacial Stage when the first Quaternary marine unit began to lie down. During this transgressive interval, three short-term environmental fluctuations of water depth can be recognized base on microfaunal data. A peak transgression of the Late Pleistocene was marked between ~35 400 and 30 900 cal a B.P.. Since about 25 900 cal a B.P., sea waters gradually retreated from the study area and the Upper Pleistocene sediments were subsequently subaerially exposed during the Last Glaciation Maximum, which is indicated by a mottled clay layer in the core. Around ~6 000 cal a B.P., the postglacial sea level rise initiated a new phase of the Holocene sedimentation. This timing was later than in other parts of the Pearl River Delta because core site is located in the Guangzhou-Panyu fault block with a higher elevation during the Early-Middle Holocene. A supralittoral-zone environment developed during the first phase of Holocene sedimentation. Since ~3 700 cal a B.P., the study area gradually changed into a tide flat environment with microfossils sporadically present.
Key words:the Pearl River Delta; foraminifers; ostracods; Late Quaternary; palaeoenvionment
中图分类号:Q913
文献标志码:A
文章编号:0529-6579(2014)06-0045-08
*收稿日期:2014-08-10
基金项目:国家自然科学基金资助项目(40872024);德国学术交流中心(DAAD)与国家留学基金委(CSC)国际合作交流资助项目